 Biologists like to classify selective events. Natural selection, artificial selection, sexual selection, kin selection, group selection and observation selection are all examples of selective categories. This post is about two more categories - self selection, and other selection. These are classification categories which mainly apply to individual decisions.
- Self selection involves decisions that affect the actor's own fitness. Positive self selection increases fitness, while negative self selection decreases it. Deciding to raise offspring is usually positive self selection. Deciding which of your shoelaces to tie first would not be self-selection - while it is a decision, it has negligible impact on fitness.
- Other selection involves decisions that affect the fitness of others. Mate selection is a well-known example of positive other selection. Selection of a rival for combat is a well-known example of negative other selection. However there are other examples of other selection which are not associated with sexual selection. Choosing a trade partner can increase others fitness of others. So can choosing who to groom, who to go hunting with or who to share food with. Nepotism counts as other selection, although it is also a type of kin selection.
Self selection and other selection are not intended to be mutually exclusive categories. Sexual species feature more other selection, as do more social species. I mentioned individual decisions above, but "individual" can be interpreted broadly to include all kinds of loosely defined groups that can be assigned fitnesses. Companies and families would be examples.
The terms can also be applied at the level of the gene. That's an interesting level because there, other selection is not "polluted" by cases of kin selection. However, it if often polluted by cases of linkage instead - genes can affect the fitness of other genes via linkage. The other selection is most useful when these "polluted" areas are not involved. I said that these forms of selection "mainly apply to individual decisions". What does it mean for a gene to "make a decision". It means it affects some behavioural outcome, relative to its alleles.
I've looked into the history of these terms a little. A common use of "self selection" is via self-selection bias. That usage doesn't seem very closely related because it makes no mention of fitness.
I've long sought better ways of explaining and teaching evolutionary theory. A recent novel approach involves evolutionary topology. One approach to applying topological ideas to evolution is to apply it to the histoy of the relationships between the agents involved. This is commonly done when constructing family trees. Here is a set of primitive operations:
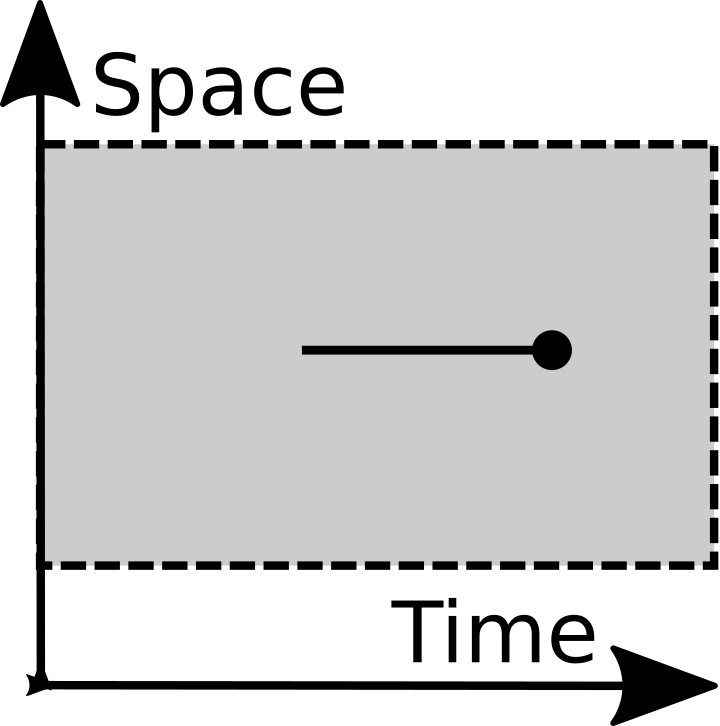 Creation |
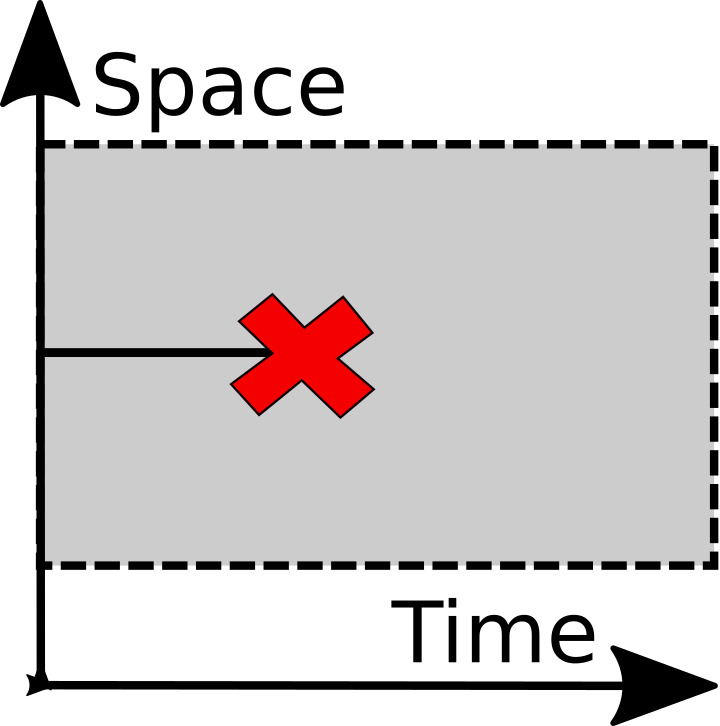 Destruction |
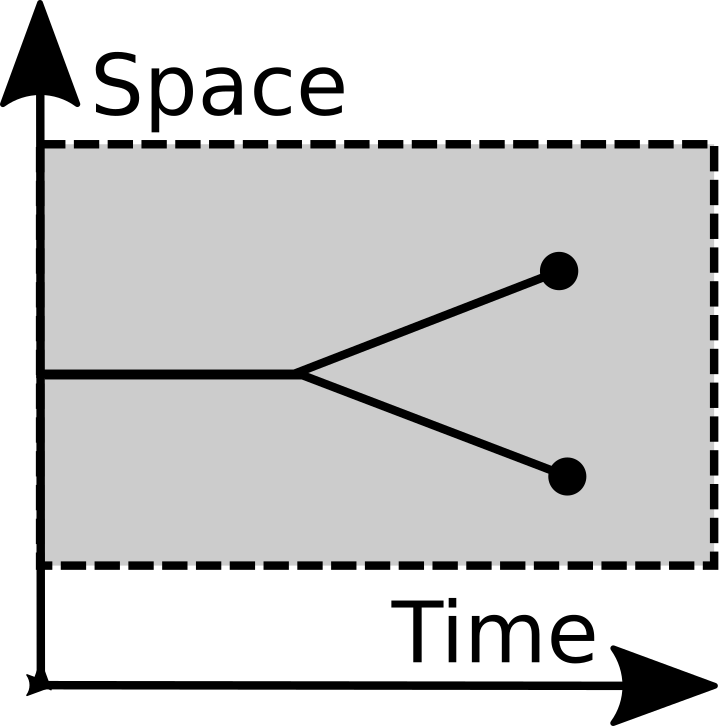 Splitting |
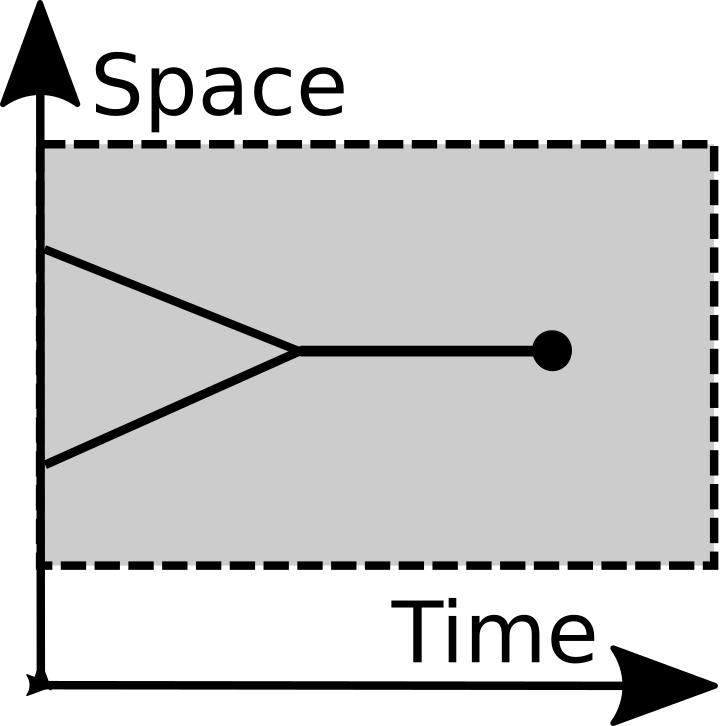 Merging |
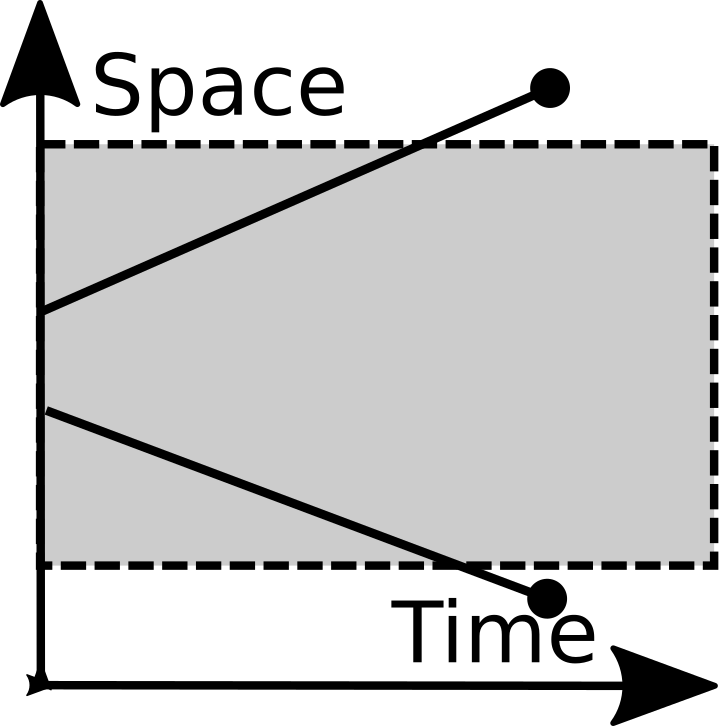 Emigration |
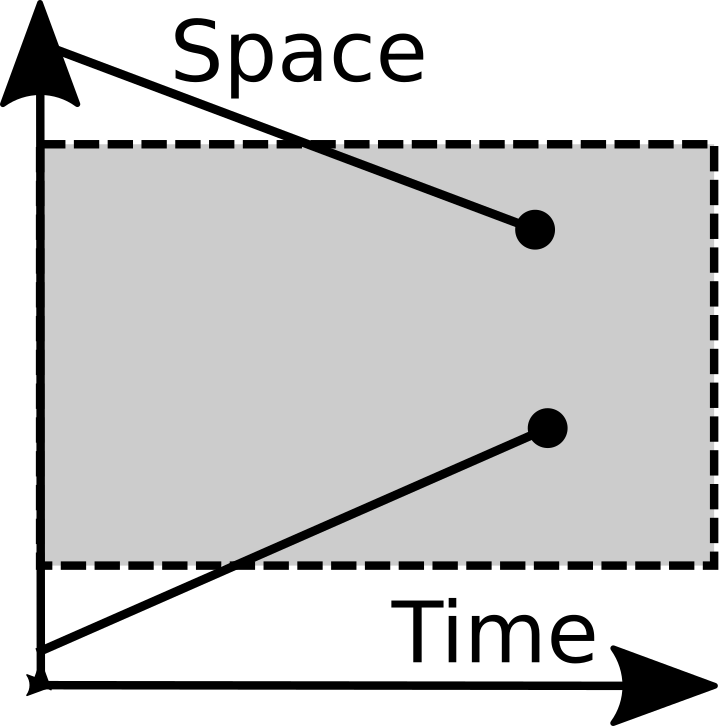 Immigration |
The operations are fairly self-explanatory, but some notes may help. To start with, this is a dualistic agent-environment framework. If it isn't clear what the agents are, that's the first thing to sort out. Agents have no properies or attributes - those would be modeled independently. Similarly there is no spatialization - again that would be modeled independently.
"Creation" refers to the construction of agents from non-agents. The most common way that agents originate is normally not "Creation" but "Splitting". An example of creation is the origin of life. However, creation can be more common than this suggests. For example as well as a first organism, there was a first ants nest and a first company.
The framework represents merging and splitting events from symbiology. Splitting is a common evolutionary primitive operation, but the significance of merging was not fully appreciated until the 1960s. Until then recombination was the only "merging" operation most evolutionists considered. Then it became seriously entertained that eucaryotic cells had symbiotic origins. While this endosymbiosis subsequently became widely-accepted science, it didn't have much effect on the foundations of evolutionary theory. Instead it was typically tacked on as an afterthought. Endosymbiosis was considered to be a rare accident during the history of life. Here, by contrast, merging is a primitive operation. It is the time reverse-operation of splitting.
Lastly immigration and emigration are included as fundamental operations. They are listed mainly because they are so important. If you are dealing with a closed system, feel free to reject them.
The framework is an alternative to the "selection-drift" framework, which seems to be one of the main ways in which the concepts of evolution are introduced to people. Selection includes both splitting and destruction, while here those are very different categories.
Historically this framework grew out of my Natural production and natural elimination. I noticed that adding more categories would help to improve the overall clarity. The concept of "evolutionary topology" is not mine. For earlier use see (for example): Topology of viral evolution (2013).
Genetic drift is a well-established idea in population genetics. It generally refers to stochastic changes in gene frequency that are not caused by selection. The term comes from nautical language: if a ship is not driven by the wind it may "drift" around, pushed hither and thither by the waves. Historically, genetic drift was not considered to be a very important force until the 1960s, when it was shown to be significant experimentally, and it was discovered that many genomes were full of useless junk DNA, which was then subject to genetic drift, resulting in molecular clocks useful for dating evolutionary divergences.
Genetic drift results in "neutral networks", whih in turn allow populations to maintain diversity that can be recruited by natural selection if the environment changes. Drift results in historical contingency and path dependence.
Complicating the definition of genetic drift is the desire to distinguish it from genetic draft. The term "genetic draft" refers to gene changes caused by linkage and selection at other loci. It is related to the concept of genetic hitchhiking. Like genetic drift, genetic draft can have a stochastic component, as a gene's neighbours vary over time. Unlike genetic drift, genetic draft can act persistently in the same direction.
Memetic drift is pretty much the exact same thing with memes instead of genes. Like gene frequencies, meme frequencies drift around in the absence of directional selection, resulting in loss of memetic diversity in small meme pools. As with genetic drift, memetic drift is a useful null hypothesis when dealing with the issue of whether an observed trait is an adaptation. As with genetic drift and genetic draft, we can distinguish between memetic drift and memetic draft.
Genetic drift is much discussed but memetic drift is rarely mentioned. I remember a good discussion of it in Kevin Layland's book "Sense and Nonsense". It is sometimes used by researchers as a null hypothesis. However, it is much less frequently mentioned than genetic drift is. This is, I believe, largely caused by cultural evolution's scientific lag. The significance of genetic drift was not recognised until the 1960s. Cultural evolution lags behind its organic counterpart in many ways, and this is plausibly one of them.
If you think about optimization frameworks in economics (one of the most advanced evolutionary social sciences) then the equivalent of genetic drift is rarely mentioned. It is the same with optimization frameworks in physics. Phyics features the "maximum entropy production principle" which explains a good deal of physical evolution. It too has concepts corresponding to genetic drift. However, it rarely mentions or makes use of these concepts. Phausibly, it is because these sciences are underdeveloped relative to evolutionary biology.
I'm generally a critic of contrasting "genetic drift" with "natural selection" (for details see here). Indeed, the definition of genetic drift is relatively complex, and it is often not a very useful scientific category, due to the practical difficulty of identifying it in any particular case. Memetic drift has much the same set of problems. However, I do, of course, acknowledge the significance of evolution by accident, and the systematic loss of diversity in small populations that arises due to factors which don't involve directional selection.
The term "memenomics" seems like an interesting fusion of "meme" and "economics". It seems to have become a brand at memenomics.com. I rather like the word but am less convinced by the associated MEMEnomics book and its "vMemes". That seems more like marketing and self-promotion than science.
I think "memenomics" should just refer to some ground in the vicinity of economic approaches to memetics and evolutionary approaches to economics. "Evolutionary economics" already has a pretty nice name, so maybe "memenomics" could be used to refer to economic approaches to memetics. A nice example of this is the well-known idea of an "attention economy" - where attention is a scarce resource that memes compete to monopolise. That's an example of applying economic thinking to memetics, but there are other ways in which economics could be applied to memetics. For one thing, attention is not the only resource that memes are interested in. They also compete for storage space, transmission bandwidth and various other resources.
 In symbiology, "transmission vectors" are the name for symbionts
that carry their partners around. So: mosquitos are "vectors"
for malaria and deer ticks are "vectors" for lyme disease.
In memetics (and genetics), it is quite common to use "vehicular" metaphors when
describing these. So, for example, we have:
The first two seem to cover many of the most significant cases. There's quite
a bit of conceptual overlap between them. Until recently I have preferred to
use the "memetic hitchhiking" terminology - largely because "genetic hitchhiking"
is well-established terminology. However, in this post I want to reexamine
the "memetic hijacking" terminology. I want to raise and address the question
of whether these concepts compete, and whether they can coexist.
What is the difference between hijacking and hitchhiking? It is partly
one of consent - a hitchhiker has permission to ride in the vehicle
while the hijacker does not. Outcomes also differ - a hitchhiker rarely
damages the vechicle or its owner, while a hijacker often does so. Another
difference is control - hitchhikers rarely alter the destination,
rarely control the vehicle and rarely eject the owner - while hijackers
fairly often do these things.
With these differences in mind, it seems fairly clear that hijacking and
hitchhiking are probably different enough concepts for
memetic hitchhiking
...and...
memetic hijacking
to coexist.
At first glance, the idea of the rider having "permission" to ride in the vehicle
seems irrelevant in the context of memes and genes. However, we can conveniently substitute
whether the guest rider is beneficial or not - on the grounds that deleterious
riders would not normally be granted permission to ride - if we "agentify" the memes or
genes involved.
This gets us on to the topic of usage in genetics. There, "genetic hitchhiking",
is standard terminology - and hardly anyone uses the term "genetic hijacking".
However if the difference between hitchhiking and hijacking is the sign of
the fitness difference the guest rider makes, then maybe geneticists should
start doing so.
As you can see, I have warmed up to the "hijacking" terminology. That the
contraction
memejacking
exists is another point in its favor in my opinion. It is true that it is a
significant problem that there's no "genejacking" - but maybe there should be.
 Evolution is notoriously a competitive business. Some adaptations have evolved for the specific purpose of doing-in competitors. There are many adaptations for male combat whose main function is doing in other males. The enlarged claws of male fiddler crabs are mainly used in combat with other males, for example. Male stag beetles also have similar competitive adaptations in the form of their claws.
Many plants do similar things. Black walnut trees load their roots and nut hulls with a toxin which seeps into the soil and kills or damages competing trees and plants. Many conifers line the ground around them with a thick blanket of needles which acts to suppress competitors. Some load the blanket with flammable resin, encouraging regular forest fires which only large, mature trees can survive.
Competitive adaptations are also common in cultural evolution. Some examples:
- "Thou shalt have no gods before me" is a famous example from Christianity.
- The Bible also features a prohibition on idolatry, with a similar intent:
You shall not make for yourself an idol, or any likeness of what is in heaven above or on the earth beneath or in the water under the earth. "You shall not worship them or serve them; for I, the LORD your God, am a jealous God, visiting the iniquity of the fathers on the children, on the third and the fourth generations of those who hate Me, but showing loving kindness to thousands, to those who love Me and keep My commandments.
- One way of exterminating the competition is killing their human hosts. Islam has plenty of examples of this with its holy war on infidels and repeated calls to violent action against unbelievers.
- American elections heavily feature negative advertising, whose sole purpose is destroying the competition. For a famous example see the daisy girl video.
- There are so-called Anti-competitive practices whose function is to eliminate competition. These are actually forms of competition in which organizations sabotage their competitors in various ways - often in the hope of eliminating the competition altogether and gaining a monopoly.
 In a 2016 post titled Shared interests of unrelated symbionts I discussed how unrelated symbionts often had shared interests, resulting in them pulling their hosts in similar directions.
A classic example of this involves promoting interactions between hosts. In the organic realm, rabies makes hosts want to bite each other while toxoplasmosis makes hosts unafraid of each other and attracted to each other's urine. In the cultural realm, missionaries seek out potential converts and teachers seek out pupils. In each case interactions between hosts are promoted by symbionts - because they need such interactions to reproduce.
Another example is reduced fertility. Many parasites compromise host fertility - probably since host reproduction uses resources which might otherwise go into symbiont reproduction. Many parasites go in for complete host castration - they are called "parasitic castrators". Many cultural symbionts also reduce host fertility - as seen in the demographic transition. Places like Japan where there are many memes have sub-replacement fertility.
This post is mainly proposing terminology. I think we should call these shared interests a "consensus". It's the consensus of the symbionts that the hosts should get out more, meet more strangers and not have kids of their own. Of course, "consensus" is not meant literally here: no-one is suggesting that the symbionts communicate via town meetings. The consensus might be different depending on which group of symbionts are under consideration. Gut bacteria might have a consensus that the host should go the the smallest room more frequently and spend more time there - while cultural symbionts might have a quite different consensus. We could call the cultural symbiont consensus the "memetic consensus" for short.
 Back in 2012 I wrote a landmark article on common descent in cultural evolution titled cultural common descent. Alas, my article included no references because - as far as I knew at the time - nobody else had seriously considered the idea. Revisiting the idea in 2017, my views have not changed very much. Nor has the situation with other researchers. Maybe my search skills are lacking, but: where is everyone?
Cultural common descent remains a useful concept which forces researchers on to the horns of a dilemma: either reject or accept common descent for life on earth. All the evidence isn't yet in - but common descent still looks like a pretty useful concept that can be applied to cultural evolution too. What that means is that researchers need to consider the possibility that memes evolved from genes or gene products.
Doesn't the "face on mars" meme disprove cultural common descent? No more than do inheritance of knowledge about gravity, water or rocks from the inorganic environment disprove common descent in the organic realm. Perhaps some will respond that of course, some memes evolved from genes or gene products. However, I think it is fair to say that this possibility is not really on the horizon of many cultural evolution researchers - and it is far-from obvious how widespread memes having purely non-memetic ancestors is in modern times. Maybe - as in the organic realm - established creatures have occupied their niches and eat them and their lunch.
IMO, what we really need at this stage is more researchers to join in. Cultural common descent has been a neglected concept for far too long now. That's rather puzzling because you might think that common descent is a core evolutionary concept and that philosophers of evolution would be eager to get their teeth into the issue. IMO, it is now time to put the concept firmly on the map.
 Johnnie Hughes once likened pioneer species colonizing a new environment to memes colonizing an infant's mind. He explained how the early species in an environment create the ecosystem for those that follow them. He then likened this to the way in which early memes create a mental environment for the more complex ones that follow them.
There's another way of looking at the educational process involving dependencies. It is widely understood that learned concepts often have prerequisites. Knowledge often depends on previous knowledge. For example, understanding written sentences depends on an understanding of the words involved which in turn depends on a knowledge of the alphabet. Knowledge can thus be pictured as an edifice in which higher structures depend on lower ones.
However, in large construction projects, scaffolding is often used. Scaffolding supports the structure while it is under construction and is then eventually removed. It seems obvious that some learning materials play the role of scaffolding in the construction of knowledge. For example, ABC books are on the bookshelves of toddlers, but not the bookshelves of adults. Adults don't need them any more.
Some concepts too get discarded during the learning process. I can clearly remember as a child thinking of my reputation as a nebulous fog that surrounded me which other minds interacted directly with. That might have been a useful concept which helped me to avoid making mistakes at the time, but I now know that it was a largely mistaken idea. Santa Claus, the tooth fairy, God, heaven and hell are all ideas which are regularly taught to children and are later discarded as the child grows up.
Educational scaffolding has been well studied by developmental psychologists since the 1950s. This Wikipedia article has more details of that.
 Scaffolding, I would argue, is an abstract engineering concept which is useful for building all kinds of structures, from buildings to scientific theories. We could have a scaffolding theory that abstracts away substrate specific details and is applicable in a wide variety of domains. It could cover issues such as the following: Scaffolding, I would argue, is an abstract engineering concept which is useful for building all kinds of structures, from buildings to scientific theories. We could have a scaffolding theory that abstracts away substrate specific details and is applicable in a wide variety of domains. It could cover issues such as the following:
- What type of scaffolding to use;
- How much scaffolding to use;
- When to add scaffolding;
- When to remove scaffolding;
- How to attach the scaffolding;
Details would no-doubt be domain specific, but we can still develop an abstract theory that is widely applicable.
Scaffolding is also a useful concept in biology. One application domain is ontogeny. The placenta is an example of developmental scaffolding that is discarded after being used. Removal of scaffolding sometimes leaves scars - and in this case, the belly button is an example of a scar marking a scaffolding attachment point that persists throughout life. A corresponding example from cultural evolution involves baking a cake. A cake tin acts as scaffolding for the cake. As with the belly button the tin leaves a scar that persists throughout the life of the cake. Another application domain is evolutionary theory. Evolution critic Michael Behe once defined the concept of "irreducible complexity" in his book Darwin's Black Box as follows:
A single system which is composed of several interacting parts that contribute to the basic function, and where the removal of any one of the parts causes the system to effectively cease functioning.
He went on to argue that "irreducibly complex" systems cannot evolve by a process involving small changes. However, of course such systems can evolve by using small changes - if they employ scaffolding. An stone arch depends on every stone: remove one stone and the arch collapses. However an arch can still be built by a gradual process of adding and removing stones. The key to construction is to use a mound of stones under the arch that supports it while it is being created. The mound is removed once the arch is complete.
For scaffolding in evolution, a lot of the engineering concerns listed above don't apply. Instead what would be useful are theories about how to identify details about missing scaffolding after it has been removed.
 Most of those who complain about gene centrism in evolutionary biology don't understand the position of the gene promoters. However, I have recently found a few who do at least address the position I hold: that genes are units of heredity and genetics is the science of heredity.
I mentioned David Dobbs's critique recently. I have also found Denis Noble has made explicit the criticism that defining a gene as a unit of heredity is unfalsifiable:
According to the original view, genes were necessarily the cause of inheritable phenotypes because that is how they were defined: as whatever in the organism is the cause of that phenotype. Johanssen even left the answer on what a gene might be vague: ‘The gene was something very uncertain, “ein Etwas” [‘anything’], with no connection to the chromosomes’ (Wanscher, 1975). Dawkins (Dawkins, 1982) also uses this ‘catch-all’ definition as ‘an inheritable unit’. It would not matter whether that was DNA or something else or any combination of factors. No experiment could disprove a ‘catch-all’ concept as anything new discovered to be included would also be welcomed as a gene. The idea becomes unfalsifiable.
This is, I think, a criticism that is very easy to refute. Science consists of, among other things, hypotheses, axioms and terminology. Hypotheses are what needs to be falsifiable. Axioms and terminology, not so much. You can criticize an axiom by saying it is false, but saying it is always true isn't much of a criticism. A similar situation applies to terminology. You can say terminology is unhelpful, unproductive, misleading and so on. However, the criticism that it is unfalsifiable cuts no ice. In science, it is hypotheses that need to be falsifiable. Unfalsifiable terminology is fine. It I say an atom is a stable mixture of protons, neutrons and electrons that can't be subdivided without changing its chemical properties, nobody can say that that's a bad definition because it is unfalsifiable. That's just a misunderstanding of how science works.
Many scientists are enthusiasts for dividing the world up into pieces, in order to study it. One of the simplest approaches to doing this involves spacetime partitioning. The partitions involved are usually disjoint. Partitioning schemes are often not unique. For example, on the left is one partitioning scheme, dividing the universe into organisms and environment, while on the right there's another partitioning scheme, dividing the universe into cells and environment.

With a perfect model of the dynamics of the contents of each partition and how they interact, different partitioning schemes would produce the same results. However, in practice models are imperfect and the partitioning scheme used can affect the results.
Spacetime partitioning schemes often work best when the partitions don't overlap. Of course, in reality organisms can have joint phenotypes - and can sometimes overlap. How can that be modeled? One approach is to use partially overlapping partitions. That generally leads to more complex and awkward models, but it is one approach. Another possibility is to partition genotypes, rather than phenotypes. Joint phenotypes are common, joint genotypes are much less common. In symbiotic relationships where joint genotypes occur, inheritance usually follows one of the partners, so even then, partitioning can be made to work reasonably well. The main problem with partitioning by genotype is that this fails to divide up phenotypes properly - they just get left out. It is a bit like hiding the problem. Sometimes you can model overlapping organisms using independent partitions. If your hosts and parasite phenotypes overlap, you can use one partition for the hosts and one for the parasites - where each is the environment of the other.
 In modern times the study of Darwinian evolution has become split across multiple academic departments. There are evolutionary biology departments, ecology departments and departments of human evolution. Other departments study evolutionary economics and evolutionary epistemology. As far as I can tell, practically nowhere studies evolutionary theory itself.
The success of the field of evolutionary biology seems to be part of the problem to me. There are a number of academics who self-identify as "evolutionary biologists". There are evolutionary biology departments at universities all over the world. It seems to me that students of evolutionary theory who confine their attention to biology are missing out on Darwinian physics, and applications of evolutionary theory outside biology. Is this just a case of specialization? or do these folk not understand that Darwinism is more broadly applicable? Experience suggests that the latter hypothesis is usually the more accurate one.
For example, Mark Ridley's "Evolution" textbook says (3rd edition, page 4):
Evolution means change in living things by descent with modification
"Living things"? Since when is evolutionary theory limited to "living things". What about Darwinian physics? To me that's a classic example of the confusion associated with evolutionary biology. Those folk think they have a monopoly on evolutionary theory. What they actually have is bad terminology which is confusing the next generation of students.
Larry Moran once wrote: "Call me an evolutionary biologist". Well, OK - but it seems like a term of abuse to me. When I refer to people as "evolutionary biologists" I am usually referring to people who don't understand the true scope of Darwin's legacy.
Superstimuli are stimuli that are larger than life. They are typically exaggerations of a particular natural stimulus. The stimulus involved could be attractive, aversive, or it could evoke other reactions - such as hunger, fear or jealousy.
Superstimulii are typically ways in which organisms manipulate other organisms. Extra-strong stimuli evoke powerful reactions - sometimes against their owner's will and best interests. This makes them useful for those who seek to manipulate others.
A picture is worth a thousand words, so without further ado, here are some superstimulii:
Superstimuli can arise in organic or cultural evolution. In the organic realm, flowers present superstimuli to insects - including genitalia mimicry, powerful scents and super-sweet nectar. Cultural evolution has led to super-sweet chocolate gateau, super satisfying donuts, super-smelly perfumes, and all manner of exaggeration of female ornaments. Pornography makes extensive use of sexual superstimuli.
Superstimuli are often used in marketing and advertising. The Daisy advertisement used a fear superstimulus relating to the end of the world for political ends. Environmental activists regularly invite people to help to save the planet. Highly attractive female models are used to sell many products.
 I've been promoting an evolution revolution over the past decade.
The main theme is an expansion of the domain of evolutionary theory. The main proposed title is Universal Darwinism.
The extent to which this is a revolution has been debated. I think it is the biggest scientific revolution I have seen and the most interesting revolution that is currently going on.
Recently, other evolution revolutionaries have been making the headlines with their own evolution revolution. In particular, I'm thinking of the extended evolutionary synthesis. Since the proponents of this recently landed an 8 million dollar Templeton Foundation grant to pursue their ideas, we will probably be hearing more from them in the future.
The proponents of the "extended evolutionary synthesis seem to want to play down the revolutionary aspects of their proposals, but their proposed revolution has a number of things in common with the ideas I promote - so it seems appropriate to compare them.
At first glance their proposals are oriented around incorporating developmental biology into evolutionary theory. I recognize that developmental biology was left out of the modern evolutionary synthesis, but I wouldn't say that the focus of Universal Darwinism was putting it back in. This difference in perspectives gives our proposals a rather different slant.
One thing they do argue for is "Inclusive inheritance". They say:
The EES recognizes that inheritance results not just from genes but also through the transmission of a wide variety of resources (epigenetic marks, antibodies, hormones, symbionts, behavior, environmental states), through which parents construct developmental environments for their offspring. In the EES, heredity and development are closely intertwined and can include all causal mechanisms by which offspring come to resemble their parents."
IMO, this is the part of their proposal that most closely resembles Universal Darwinism. Universal Darwinism points out that copying operations are ubiquitous in nature, and are not confined to biology. High fidelity copying is also common. My main comment on their presentation is that it clearly does not go far enough. In particular, I see no sign of Darwinian physics in their presentation.
To their credit the proponents do extend inheritance beyond DNA and culture. They criticize 'dual inheritance' models of cultural evolution as ignoring other kinds of environmental inheritance and propose their own 'triple inheritance' models which don't have this defect. This material is all good and correct.
In Universal Darwinism, it isn't just inheritance that is "extended". It's all the operations found in more conventional evolutionary theory: copying, mutation, selection, filtering, sorting and merging. Do the proponents of the extended evolutionary synthesis recognize this? To some extent they do. Their presentation of this idea seems a bit odd to me, though. What they emphasize is what they call "multiple routes to the adaptive fit between organisms and environments". The basic idea here is that rather than just organisms adapting to environments, organisms change environments to suit themselves. Since they have organisms selecting environments as well as environments selecting organisms, this seems like a bit of an expansion of selection - compared with the conventional role of selection in evolutionary theory. However, this idea too seems watered down compared to the presentation typically found in Universal Darwinism.
I've previously commented about their terminology. They use the term "niche construction" use a lot, but have a confusing and counter-intuitive definition of it. According to them "niche construction" involves any change made by an organism to its environment. That mixes together niche creation and niche destruction. In my opinion, referring to destructive operations as "construction" gets confusing fast. I call their concept "environmental modification by organisms". I don't think it is important enough to deserve a snappier title. Things like "environmental modification", "niche creation" and "niche destruction" seem like better terminology to me.
They frequently use the term ecological inheritance. By contrast I more frequently use the term "environmental inheritance". I don't pretend these terms are synonyms, but they do represent competing terminology. Ecological inheritance seems like an umbrella term that includes organic inheritance. Environmental inheritance excludes organic inheritance. So we have Ecological inheritance = Organic inheritance + Environmental inheritance. It's nice to have an umbrella term, but IMO, usually what you want is something that contrasts with organic inheritance - not something that includes it.
I'm also concerned that their more revolutionary content is mixed up with a lot of more conventional material that will be regarded as being old hat.
Maybe there are some good things in their proposal that I'm currently missing, but my impression is that this is a watered down and inferior evolution revolution - when compared to the Universal Darwinism that I have been promoting. In fact, my reaction to their proposals has been similar to when I first read about Boyd and Richerson's version of cultural evolution. I felt as though the revolutionary content in memetics had been watered-down and stripped out, leaving a rather dry and boring husk. Perhaps watered-down content is all that academia is likely to stomach - but I want evolutionary theory to skip ahead a bit. The evolution revolution has already been going on for 150 years. How much longer is it going to take?
References
 I have long had an entry for "semes" on my meme synonyms page. However, I only recently read up about them. Barry Hewlett, a colleague of Cavalli-Sforza seems to have been one of the main proponents. Here's how one of Barry's papers introduces them:
Following Cavalli-Sforza, we call these units “semes” rather than “memes” (Dawkins 1976, Durham 1991, Boyd and Richerson 1985) because “seme” comes from “sign” and emphasizes the symbolic nature of culture.
Semes were apparently first mentioned in a 1970 book by Roland Barthes called "S/Z" - a three hundred page dissection of a short story written in French.
However, they were subsequently adopted by Cavalli-Sforza, Marc Feldman and others. Feldman explains the term in a 2007 presentation here. He presents "semes" as the cultural equivalent of "genes".
Marcus Feldman said in 2014 (16 minutes in):
I call them semes because I don't like memes, memes gives some impression of biology which I do not want to do and semes comes from semiotics.
Of course, culture is part of biology, so biologically-inspired terminology is completely appropriate.
I think "semes" are more-or-less dead terminology now. Not all culture is symbolic, so the death of "semes" seems to be no great loss. However, IMO, it is fascinating historical tit-bit that Cavalli-Sforza and Marc Feldman join all the other students of cultural evolution that publicly toyed around with "-eme" words.
 The concept of natural selection part of the core of Darwinian evolutionary theory. However
in my opinion, it has become muddied by definitional issues. One issue is the term has come
to cover a mixture of processes. It covers both death and reproduction. Another is that some
biologists decided they wanted to use the definition of natural selection to distinguish
between changes likely to produce adaptation and those likely to destroy adaptations -
creating a distinction between selection and drift. I'm not too happy about the results of that effort.
I've previously argued for a conceptual breakdown of natural selection into components representing
death and reproduction: natural
production and natural elimination. The rest of this post describes some orthogonal concepts.
The term "selection" commonly refers to a choice between alternatives - and the term
"natural selection" was coined to indicate that the chooser didn't have to be a breeder, or even an agent: nature
could choose. However, IMO, things went rather downhill for natural selection after that.
After a while, biologists started to use the term "subset selection" to refer to this simple
choosing but they decided that this wasn't much use when considering how a sexual population
produced the next generation, since that process also involved transformational changes. The
conception of selection was enlarged to handle this more complicated case. That was, I think
a big mistake.
Scientists outside biology have another word for "subset selection". They call it "filtering".
Filtering is an important process which explains a lot about adaptation in nature.
Filtering applies to organisms when they die. Filters may also be applied while choosing mates,
or during sperm competition. However, though filtering is an important concept, it doesn't
explain everything.
In computer science, filtering is often combined with sorting. Sorting is a common
and important process in nature. Rocks are sorted on beaches and molecules are
sorted in the atmosphere. I think we can get some clarity back by using the
terms "natural filtering" and "natural sorting" instead of "natural selection".
I have previously written
a whole article about natural sorting.
Unfortunately, sometimes explanations of natural phenomena in terms of filtering and sorting
can also become contrived. One problem is that filtering is a binary process: either items make
it through the filter or they don't. Filtering is a reasonable metaphor for life or death situations.
However it seems less applicable to other aspects of reproductive success. Offspring counts are still
somewhat discrete, and it could be argued that they consist of a series of selection events.
Resource acquisition can be treated in the same way: either an item of resources is obtained
or it isn't. However, here it looks as though we are approximating a continuous
function with a series of discrete steps. That kind-of works - but sometimes, it is just
contrived and pointless: shoehorning the phenomena into the model.
When phenomena appear continuous, using a continuous model makes sense. My proposal for a
continuous modeling framework involves the common concepts of "reward" and "punishment".
The idea is that events can have positive or negative on fitness. Positive impacts are rewards
and negative impacts are punishments. Other consequences of events are deliberately neglected -
for the sake of keeping things simple. As with natural selection, we can have agent-free
versions: natural reward and natural punishment.
Reward and punishment are common consequences of actions by agents. However here it is
important to remember that natural rewards and natural punishments may happen without
regard to the agents previous actions. There need not be any attempt reinforcing of
behaviour going on. Nature may be capricious.
In standard evolutionary theory, natural reward and natural punishment are currently
most frequently referred to as favorable and unfavorable selection pressures. What
I'm proposing here are new names for these existing concepts.
Reward and punishment are concepts from psychology. This is one of the expected
application domains. Rewards lead to reinforcement and reproductive success of
patterns af various levels in the brain, from synapses to axon firing patterns.
Similarly punishment results in weakening and destruction of such patterns. In
other words, use of these terms isn't intended as a metaphor, reward and punishment
in psychology would be a straightforwards applications of these evolutionary concepts.
 Looking at what the internet thinks about NLP (Neuro-linguistic programming) suggests that it has decided that NLP is pseudoscience.
NLP was a bit pretentious - by putting the term "neuro" in its name - but we could have lived with that. It is true that humans are programmable animals - and that they can be programmed using language. This is fairly obviously a topic for scientific study.
It seems a bit unfortunate that NLP didn't make more headway. If NLP fades away there will be a bit of a void in the niche it once occupied. I think we should probably hang on to the "linguistic programming" part. I can't think of any more appropriate terminology. We do have the well-established term "suggestion" - but "linguistic programming" is specifically to do with language, and not all "linguistic programming" comes in the form of suggestions.
The problem I see with "linguistic programming" is that you can program both brains and computers with language - but these are pretty different topics, so there is not all that much need for an umbrella term.
The most obvious alternative to this path that I see is to try and hang on to NLP - and turn it into a respectable topic. I'm sure that that path will have some advocates. I'm not sure at this stage that this plan would be effective.
 If cultures are composed of cultural organisms, what is their anatomy like, and how can they be dissected?
One of the first meme enthusiasts to propose a way of subdividing cultural entities was Douglas Hofstadter, who promoted an "anatomical breakdown" involving the concepts of "bait" and "hook". For more details, see the article: Douglas Hofstadter's contribution to memetics.
In his 2004 book, Ely Asher proposed another "anatomical breakdown" - involving "anchors", "payloads" and "carriers".
The concepts of "promises" "threats", "rewards" and "punishment" also seem appropriate as common components of many cultural creatures. Emotions are often involved in motivating humans to propagate memes, and this basic Pavlovian approach is one of the most common ones.
There's also the genotype/phenotype split to consider. In some cultural creatures there's a well-defined split between what is inherited and its associated products. Such cases typically involve an irreversible developmental program. Think of a cake and its recipe, for example - or server-side software and the web sites they produce.
I think it would be fair to say that cultural creatures come in many varieties, and that it makes sense for some anatomical structures to be domain specific. To give my usual example of different species of culture, FORTRAN programs and origami patterns need not have the same anatomical structures. However, having said that, the search for common anatomical structures in cultural creatures seems like an important one.
In some cases we can classify cultural organs as follows:
- Sensory organs;
- Motor organs;
- Information processing organs;
- Resource processing organs;
Think of robots, for example, for a place where such classification is appropriate. This path gets us into territory already explored by cybernetics and systems theory.

I'm often on the lookout for opportunities to get memetics-related content to go viral on the internet.
One potentially explosive topic is memetic hitchhiking. Memetic hitchhiking
and memetic linkage are important topics for understanding memetics, but so far they have poor coverage in academia and online. I seem to be the main promotor of these concepts.
One thing I figure I can do to help is to make the proposed terminology as polished as possible. In my original memetic hitchhiking article I used the term 'payload' to describe content
that spread via memetic linkage with another meme. That seems reasonable if the context is already memetic hitchhiking - but otherwise it is too vague. Enter hitchhiking memes:
"Hitchhiking meme" definition: a meme which spreads primarily via memetic linkage to another meme.
Meme-savvy marketers are probably the #1 audience for the memetic hitchhiking concept. It is a common marketing technique to link a brand or product with some viral content to help distribute it. The basic idea of memetic hitchhiking is pretty obvious, but the terminology to describe it scientifically is not getting used. My hope is that the idea of hitchhiking memes will give memetic hitchhiking a boost.
 In 2014 I declared: I Am A Social Darwinist.
I was protesting against the hijacking of the basic term by critics. I figure that I am not the only Social Darwinist out there - dozens of scientists appear to me to fit the description. However, I notice that few seem to have 'come out' and publicly declared themselves to be social Darwinists. In the hope of encouraging them I invite them to take the Social Darwinist pledge - and publicly declare themselves to be Social Darwinists. A cadre of proud social Darwinists seems like the best way to combat the stigma associated with the term by critics. #socialdarwinism
|
|


























