We now at last have a significant academic literature on meme-gene coevolution. However few seem to have considered the dynamics of the meme-gene-queme coevolution that can be expected as a result of considering quantum Darwinism in the brain. This blog post is a brief attempt to share my thoughts on the topic.
The first thing to say is that it isn't just memes genes and quemes - Darwinian dymanics arise on multiple levels within the brain, for, for example, signals in the brain are copied whenever an axon divides, and are subect to selection and variation - producing a kind of neuronal spike Darwinism. Another type of Darwinian dynamics in the brain arises as a result of competition for resources between branching axon and dendrite tips. ideas are also copied with variation and selection within the brain - including ideas that don't normally qualify as memes because they were not the product of social learning.
One way in which we can expect the dynamics to differ from meme-gene coevolution is that culture is new on the scene, while the other kinds of psychological and neurological Darwinism have been going on for many millions of years. There will have been more time for the genes to adapt and reach a steady state equalibrium with these other Darwinian processes - while meme-gene coevolution is clearly out of balance and is still shifting.
An important way to understand the results of evolutionary processes is to consider their optimization targets. When there's coevolution there are usually multiple optimization targets, and one needs to understand how they interact by considering the power and speed of the optimization processes involved. Quantum Darwinism looks as though it could be fast, which means that we should take it seriously. Assuming that we reject Copenhagen-style versions of Quantum Darwinism in which branches of the wavefunction collapse and die, quantum Darwinism is a kind of splitting only, quasi-Darwinism - where differential reproductive succees in important while differential death is not. With this perspective in mind, the "goal" of quantum evolution appears to be to put us in the most split (and most splitting) worlds. One way to understand the implications of this is to take a thermodynamic perspective. World splitting is populatly associated with irreversible thermodynamic effects. What that means is that quantum Darwinism can be expected to behave like other kinds of Darwinism - in terms of maximizing entropy production.
I think this thermodynamic perspective helps get a handle on the significance of quantum Darwinism in the brain. If the brain ran hot, there would be lots of scope for quantum Darwinism in the brain, while if it runs cool, there's less scope for quantum Darwinism to operate. Most agree that the brain is on the cool side - considering what it is doing.
I think that genes are likely to be optimizing for cool brains, and brains that optimise for gene-coded functions. This may often pit them against quantum Darwinism in the brain. A cool brain is good news for quantum computation theories of mental function (fewer thermodynamic irreversible events means less chance of decoherence) - although those look implausible to me on other grounds. However a cool brain doesn't help the argument for quantum Darwinism being important in the brain.
Evolutionary processes liek to "harness" each other, to bend their optimization targets towards each other. Because quantum Darwinism in the brain has coevolved for millions of years with the genes, they have had a long time to find ways to harness the power of quantum Darwinism. However, the classical way for one evolutionary process to harness another one is by altering its fitness function. The genes might find it hard to affect the fitness function of quantum Darwinism since that is tied up with fundamental physics. That is going to make harnessing its effects more challenging. Another potential way for one evolutionary process to harness the effects of another one is by influencing the variants that it chooses between. However, this mechanism seems weaker and less useful.
My conclusions here are pretty tentative, but the picture I am seeing here is that the brain might not be able to make much use of quantum Darwinism because it is an alien selection process whose optimization target can't easily be controlled. In which case, the brain might be best off attempting to minimize its influence. This would be a rather boring conclusion. Mutualism and harnessing would be a much more interesting result. However, I stress again that it is somewhat uncertain. Maybe the brain can make some use of the power of quantum Darwinism by influencing the things it selects between. Or maybe evolution is smarter than I am and has found ways to make use of it that I haven't thought of.
Here's a draft of a book chapter I have written on the topic of observer and observation evolution.
Introduction
One of the places where mainstream physics has come closest to embracing Darwinism involves the role of
observers. Physicists identified the role selection of observers plays, and identified it as the cause of goodness
of fit between man and his environment - in the form of life-friendly physical laws and a stable planetary home.
History
Historically, the evolution of observers was first studied by a physicist called Brandon Carter
in the 1970s. One of the ideas he came up was was that physical constants having values that
made the universe habitable was not due to chance or an intelligent designer. It was no accident
that we observed a universe with life-friendly physical laws, since it could not be otherwise:
any observers in universes with physical laws that were not life friendly would rapidly perish.
This idea was christened "the anthropic principle" and it was contrasted with the Copernican
principle - which states that we are not in a special place in the universe. According to the
anthropic principle we are in a very unusual place in the universe - one suitable for the
evolution of humans over billions of years.
The result of the need for life-friendly physical laws is an adaptive fit between the
universe and living systems inside it. The physicists didn't describe what they had
discovered as an "adaptive" fit. Instead they said that the universe was 'fine tuned'
for life. Most of the physicists involved didn't seem to link these ideas to evolutionary theory.
Instead they seemed to consider it to be an entirely new area of science which they had discovered,
one that could explain the appearance of design without invoking a designer. They did, however
use the terminology of 'selection' to describe their findings - mirroring the terminology used in
evolutionary theory. Retrospectively, it seems obvious that they were just applying Darwin's
discovery to human observers.
Observation selection
A subsequent development was the discovery that a similar idea could be applied to observations,
as well as to observers. Observations may be filtered in a number of ways - both before and after arriving
at the senses. For example, publication bias filters information before it reaches an observer. An
observer's expectations and preconceptions might then go on to filter information further before
it reaches consciousness.
Observation of the observable
From one perspective, observation selection is one type of selection in the nervous system among
many. Filters also control whether information is stored, when it is retrieved, and
when it is forgotten. Still more filters are applied to ideas, action plans and motor outputs. However,
if you look at the situation another way, observations seem to be fundamental - since all
knowledge gains are made through observation. Observations are the basis of everything an organism
knows. Everything else consists of inferences derived from observations. This gives observations a
primary status. For example, if a fruit fly in an scientists's experiment dies, that's a case of natural
selection. However the scientist doesn't learn about it until an observation is made. We can say that
survival of the fittest is a special case of observation of the observable. This results in a reformulation
of evolutionary theory puts observers at the heart of the theory. This broadly mirrors the changes in
physics that happened at the turn of the last century, when it was discovered that observers played a
surprisingly central role in physics.
Observation reproduction
Selection is only part of Darwinism. That many observers reproduce is perhaps too obvious to mention.
However, observation reproduction merits a few comments. Behavioral reproduction is ubiquitous in
human cultural transmission. It is widely agreed that humans often copy the behavior of other humans
via behavioral imitation. However, behavior is only one side of behavioral imitation. To be copied, behaviors
have to also be observed. Observations reproduce during this process just as behaviors do. To give an
example of an observation that catalyzes its own reproduction consider the observation of yourself,
snorting cocaine. Such an observation is often followed by more similar observations. This is a simple
case of observation reproduction.
Beyond survival and reproduction
Observation selection illustrates how Darwinian dynamics can involve more than survival and reproduction.
Observation selection also filters out things that are hidden from the observer. This allows evolutionary theory
to be applied to cases where the observation of entities is based on their visibility - rather than because they
and their ancestors survived and reproduce. Survival and reproduction are important determinants of what
we observe, but they are not the only factors involved.
Multiverse
The idea that the visible universe is the product of selection effects suggests that the
visible universe is part of a multiverse. It which case it would be helpful to know the size
of the multiverse - and which parameters are free to vary in it. Alas we can only observe our small
corner of the multiverse. This leads to a difficult inductive inference problem with very little
data to go on. The concept of a "reference class" is sometimes used to denote the set of objects
being selected from. When dealing with other possible worlds it isn't always clear what the
set of worlds being selected from consists of - since we only see one world.
Brandon Carter's Ultra-Darwinism
One of the physicists who did recognize links to Darwinism was the originator the idea in the first
place: Brandon Carter. He wrote in 1992 that: "anthropic selection should be considered as an adjunct
to ordinary natural selection". He proposed that the union of anthropic selection and natural selection
be called 'Ultra Darwinism'. That's pretty much the same thing that I am saying - except that Brandon
used a different name. However, I would emphasize that the topic is mostly just applying basic
Darwinian principles to observers and observations. There are a few other topics involved too -
for example, the maximum entropy principle is used to handle ignorance. However, this is mostly
ordinary Darwinism applied to observers and observations.
Terminology
The term "anthropic" turns out to be rather unfortunate. The "anthro-" prefix means:
"man", but the basic idea can easily be generalized to cover animals, plants and machines. The
human-based version of the idea seems anthropocentric to the point of being unscientific.
I came up with the idea of branch tip evolution in 2012 by thinking about positional inheritance. Probably the simplest way to understand it is by thinking about the evolution of buds on tree branches. Buds reproduce and they undergo selection. The result is often an adaptive fit between the shape of the tree and its environment. For example, some buds may be in shade under a bridge while others may overhang a busy road. Selection affects the growth and reproduction of the buds, resulting in an adaptive fit between the shape of the tree and its environment.
Copying with selection resulting in adaptive fitness are the hallmarks of Darwinism. However this was not a form of Darwinism based on DNA genes. The example holds up even if all the tree buds involved are precisely identical in terms of their DNA.
A kind of Darwinism of branch tips can be useful in explaining a wide range of tree-shaped structures in nature. In the organic realm, there are branches, roots, corals, circulatory systems, respiratory systems, and branching axons and dentrites. Inorganic tree-shaped systems include electrical discharges, propagating cracks, crystal growth, and drainage basins.
Many models of these types of inorganic system take a functional approach to them - for example, saying that drainage basins form efficient structures for rapidly removing water from landscapes - of that they maximize the production of entropy. From an evolutionary perspective, such models are all very well, but they are all to do with adaptive function. Adaptation is part of evolutionary theory - but it also has another side: path dependence, or historical contingency. Evolutionary theory provides a rationale for adopting a functional perspective in the first place, and it also helps to explain cases where there are deviations from what strict functionalism might predict.
While branch tip evolution is an excellent and important model of many physical systems, it has limitations. In real trees, there is cell reproduction within the branches as well as at the branch tips. Also, there are other cases where branch dynamics are important - i.e. when not all the action in the system is taking place at the branch tips. Finally, real branch tips can sometimes shrink as well as grow. In such cases, the analogy between the real tree and a family tree starts to break down. More sophisticated models involving graph evolution may be a better fit for such cases.
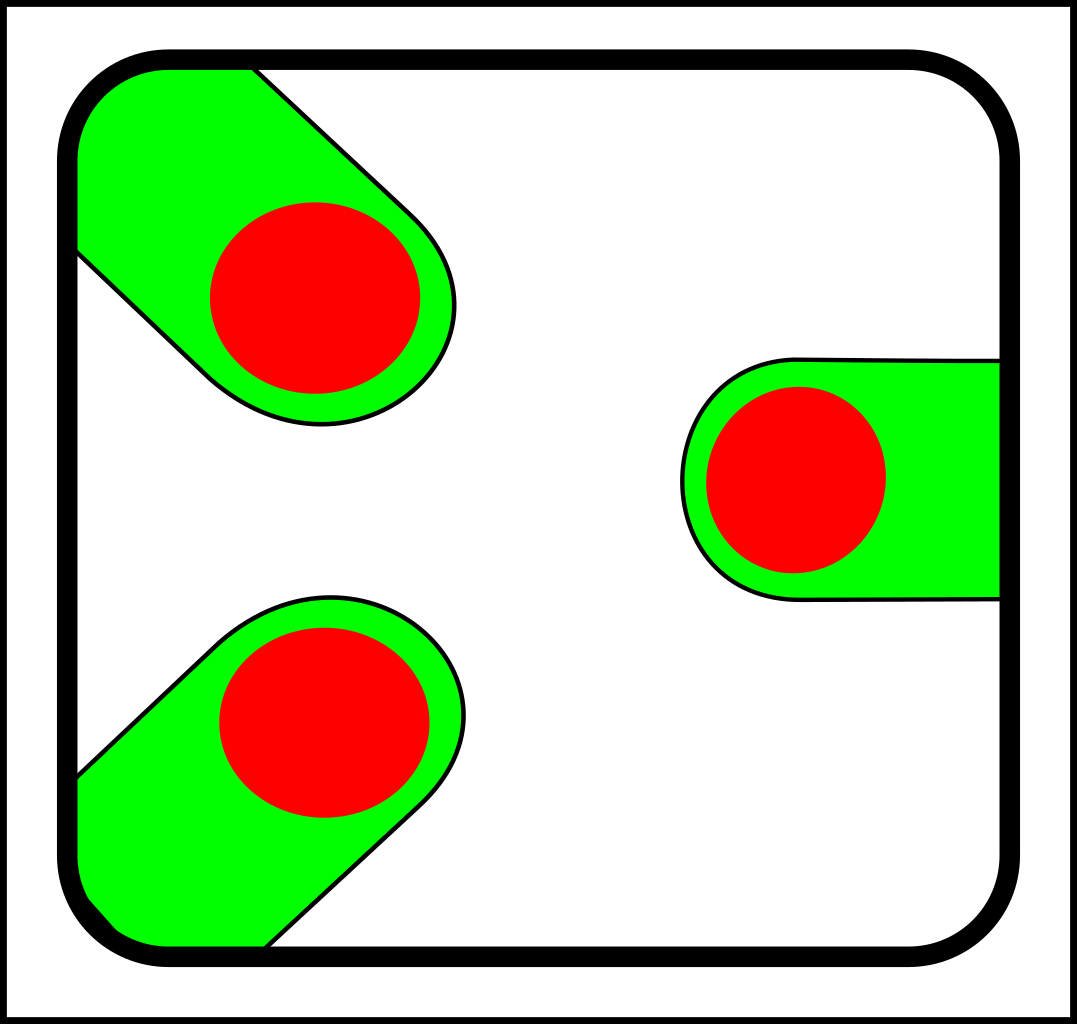
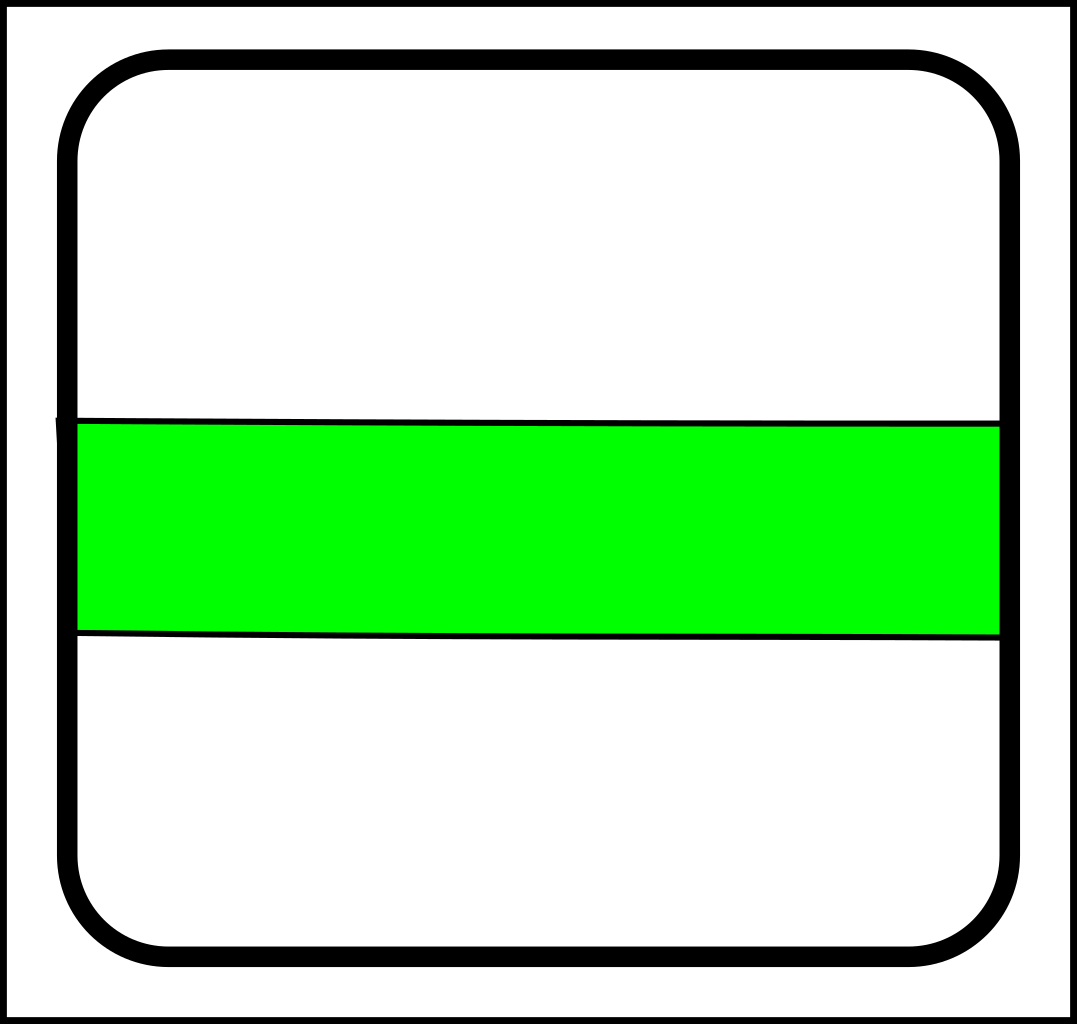
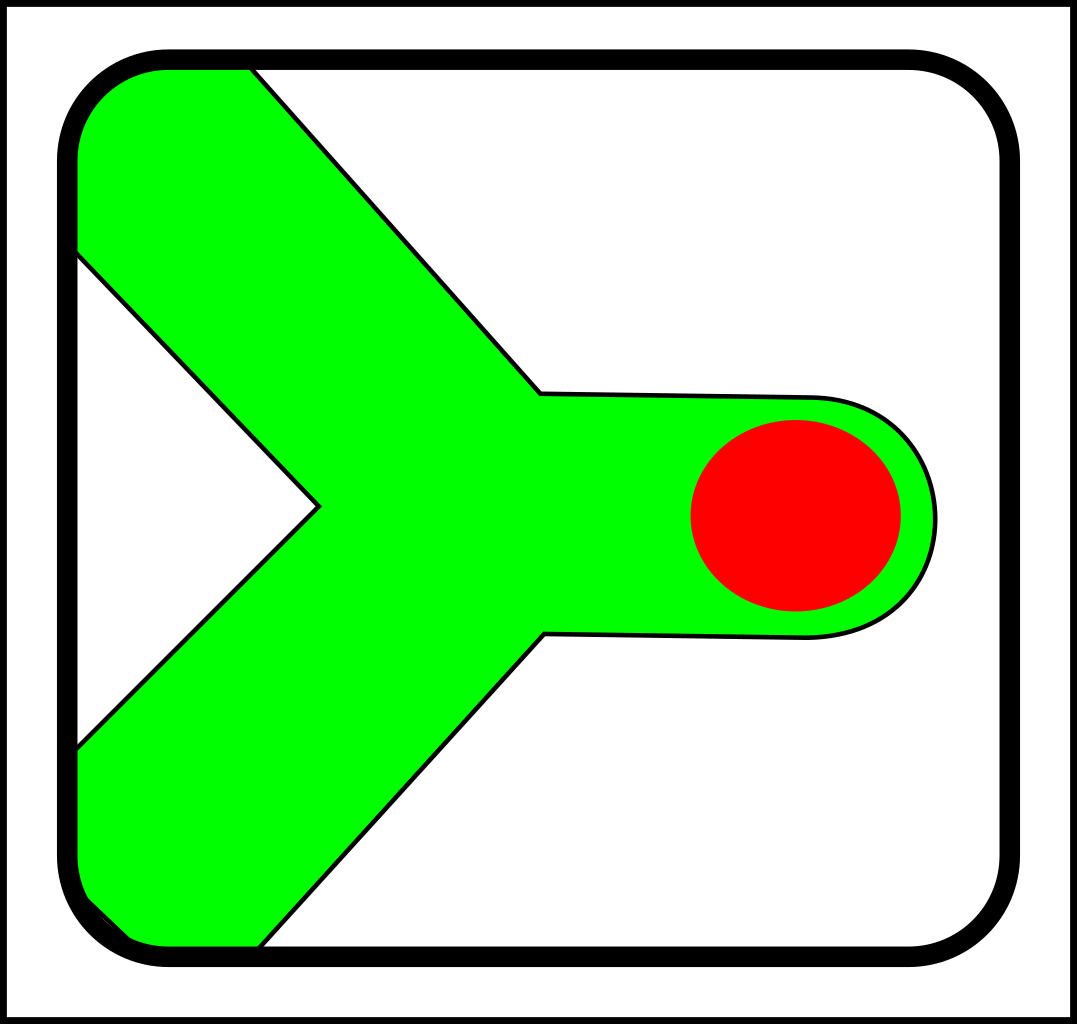
Through the first half of the 20th century evolution was mostly the science of selection and splitting. To illustrate with diagrams, a simple selection operation looks like this:
->
->
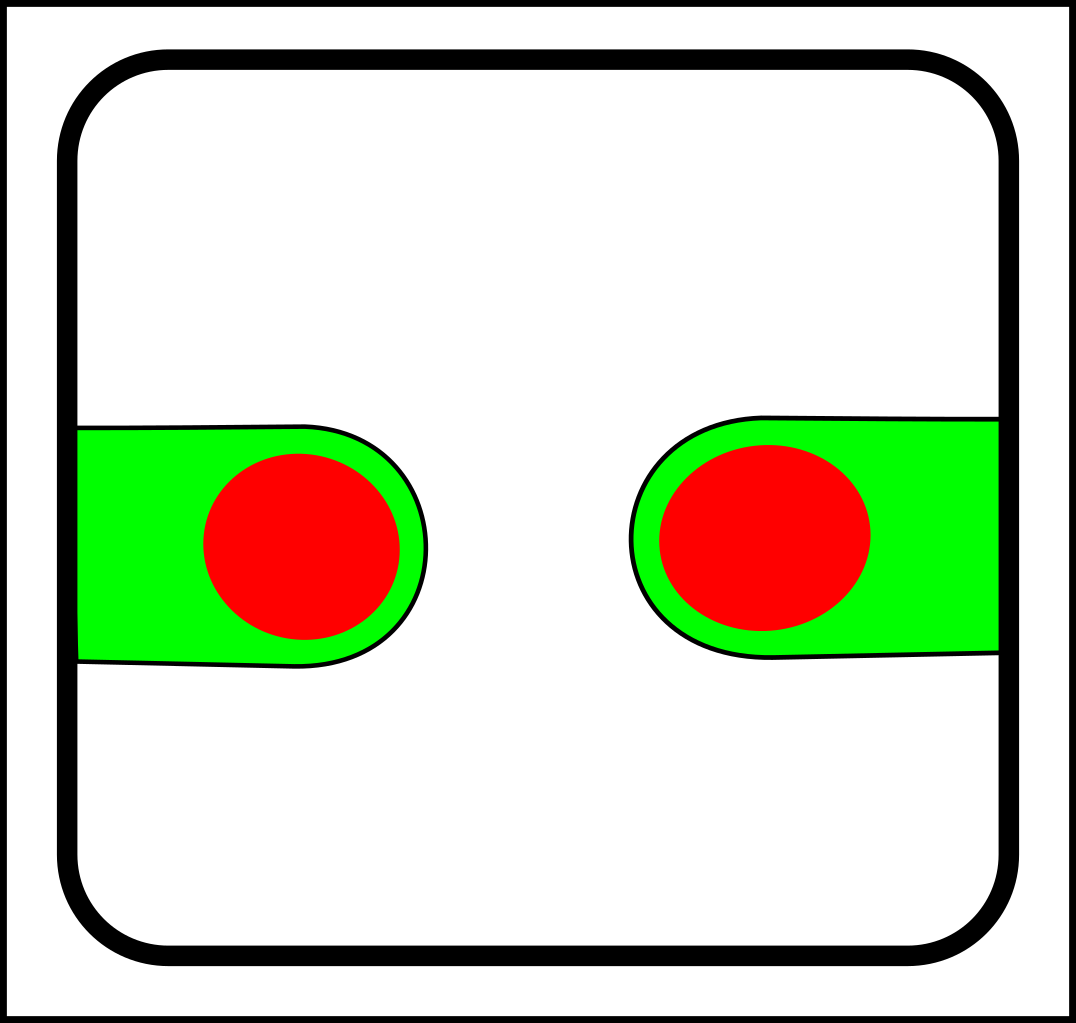
...and splitting looks like this:
->
->
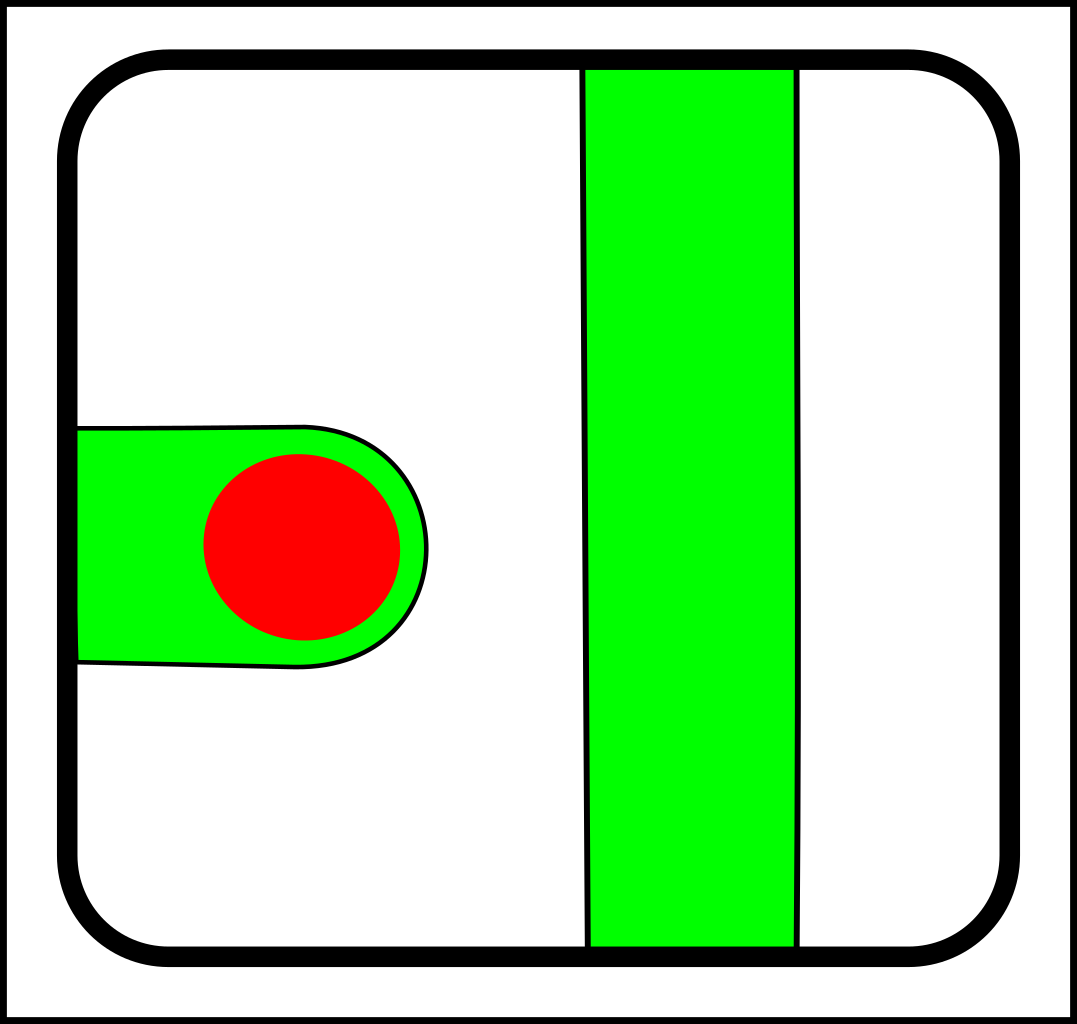
Then in the 1960s, symbiosis and sex started to look as though they needed another fundamental operation - merging - which looks like this:
->
->
These are the basic operations in modern evolutionary theory. As you can see from the diagrams, these basic operations are particulate - in that they operate on discrete, particle-like entities.
I'm thinking that a node-and-link scheme would be a better match for the evolution of brains, tree branches, circulatory systems, immune systems, respiratory systems - and so on.
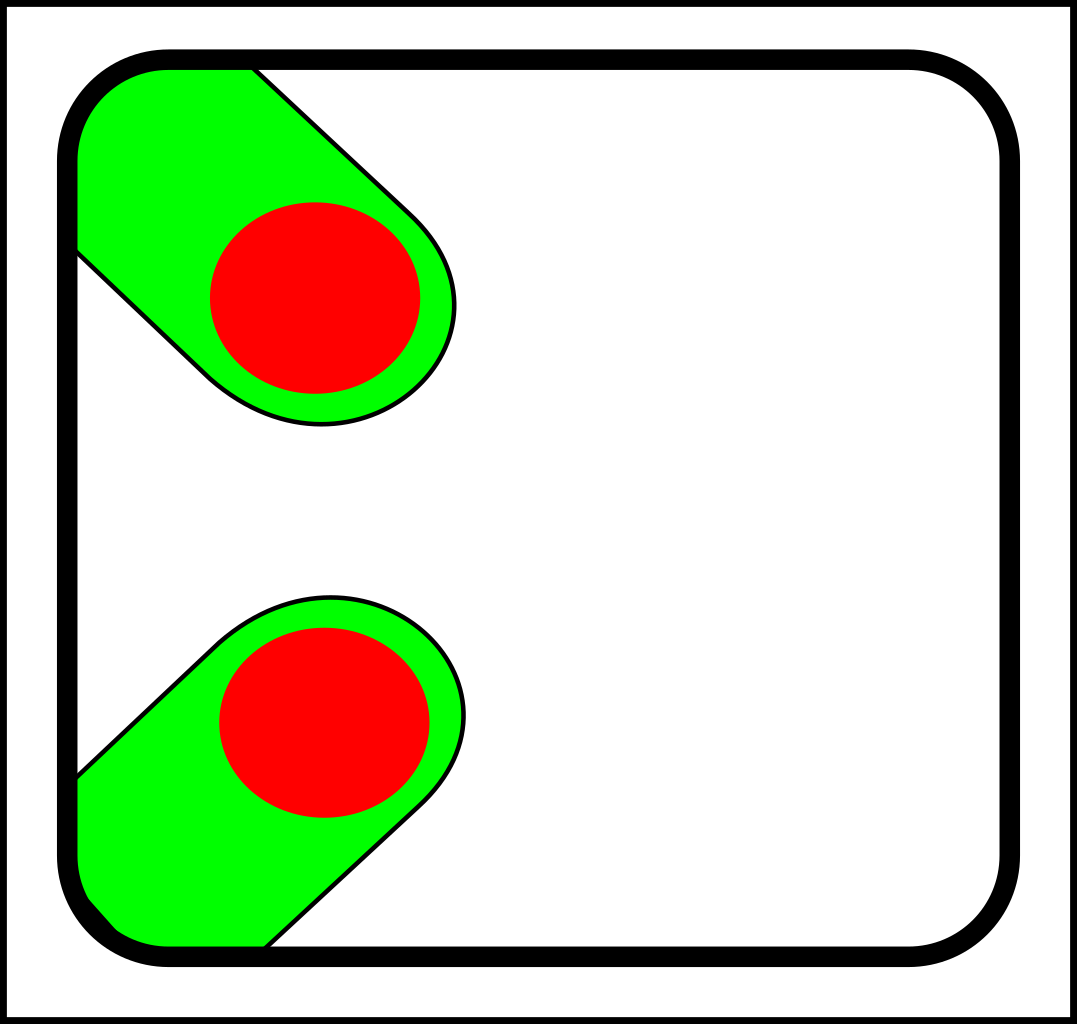
At first glance, this might seem like a case of adding operations that create and destroy links between nodes. That might make sense in computer science, but in nature, links form gradually and a more gradualistic scheme seems more in tune with existing evolutionary theory. Here are the splitting operations:
->
->
->
->
->
->
...and here are the corresponding joining operations:
->
->
->
->
->
->
For the sake of simplicity, I've only shown diagrams for nodes with fewer than four links. I've also
omitted selection events - i.e cases where elements (nodes or links) are simply destroyed.
I should probably say a bit more about the motivation for such a scheme. In biology, tree and graph evolution
is often reducible to particle evolution - since the components involved are often composed of particulate cells.
However, this doesn't work so well for brain evolution - since axons and dentrites are so link-like.
It's often possible to model branch tip evolution using conventional Darwinian models which treat branch tips as
individuals. This works quite well in many of the types of inanimate systems considered by Darwinian physics, for example. However this approach works best when flow into the resulting tree-like structures is time-like - i.e. unidirectional and at a constant speed. If resources can flow back out of unpromising roots or newly-shaded branches, conventional Darwinian models based on a population of branch tips do not work so well. Then, more sophisticated models can be useful.
There is a field called evolutionary graph theory -
but that seems to be about something rather different from the topic of this page.
Darwinism's application domain expanded in the 20th century from its traditional
area - of the evolution of DNA-based organisms. Despite vociferous objections from some anthropologists and philosophers, it is now widely recognized to cover a wide variety of cultural phenomena - including science, technology, language and religion.
Paralleling the rise of Darwinian cultural evolution, Darwinism was also applied
to the development of organisms. First, evolution within the immune system was
observed - and then various evolutionary models of other developmental stages
were found to be useful. Multi-cellular organisms can be usefully regarded as
populations of cells which themselves evolve over the lifetime of the organism
they are part of. Darwinian
models of psychological evolution were also developed.
Skinner's model of learning, for example, was explicitly Darwinian.
Gerald Edelman was involved in both the immune and neural breakthroughs.
These expansions of the application domain of Darwinian evolution led to a
back-to-basics study of Darwinism and its limits. These studies suggest that
Darwinism has applications well beyond biology.
Physics has long laid claim to observation selection effects - though the
main students of selection are those studying evolutionary biology. However,
it was additionally found that
many simple physical systems can be modeled
using traditional Darwinian models - based on copying with variation and
selection. It turns out that the tree-shaped patterns found in electrical
discharges, propagating cracks, and fractal drainage systems are all composed
of family trees - representing patterns of descent which are subject to
variation and selection. This can be seen in
slow motion images of these
phenomena.
One of the key concepts in understanding these systems is
positional inheritance -
or, more generally, spatio-temporal inheritance. When entities split, the offspring
often inherit a variety of properties from their parents. One set of properties
that are regularly inherited with high fidelity are spatio-temporal coordinates.
Other properties are also regularly inherited. Velocity, charge, temperature
and chemical composition are other examples.
In practice, many variables correlate with position. A splitting pebble inherits its
parent's position, but it also inherits its acidity level, humidity, temperature, and
many other parameters from its parent's ecosystem. This is due to
the smoothness of nature.
Nature's smoothness helps to ensure that fitnesses are inherited - which is one of the requirements for adaptive evolution.
The upshot of all this is that we can use Darwinian models to study simple
physical systems, and the concepts of adaptive fit and fitness are applicable.
The resulting field of study is known as
Darwinian physics, which is considered
to be part of Universal Darwinism.
Physicists have long realized that physical laws might cover these types of
complex systems - and various models of them have been built. Often these
models have then been exported to biology. Fractals, reaction diffusion
systems and diffusion limited aggregation are examples of physics being
applied to biology. However, physicists seem to have been reluctant to
look to the fundamental principles of biology and see how they apply to
physics. The thinking seems to have been that Darwinian principles
only apply to living systems - and that the high fidelity copying required
is rare elsewhere. On this line of thinking, biologists have Darwinism covered.
In fact copying with variation and selection are ubiquitous in nature.
Copying takes place whenever information at one location spreads to
multiple locations. This can be formalized in terms of Shannon's concept
of mutual information. The process is a common one. It happens when waves
radiate. It happens when sunlight hits dust. Selection is similarly ubiquitous.
In its simplest form, selection involves choosing a subset of items
from a set. That happens all the time - and isn't confined to the
death of living things or to choosing mates. When rocks erode, some
of them collapse into the water, while others do not. Some of the Sun's
photons hit planets, while others do not. These can usefully be
interpreted as cases of natural selection.
Many simple physical systems behave as goal-directed fitness
optimizers -
in the same way as biological systems do. They can be used to solve optimization
problems - much as genetic algorithms can. Examples of this are the way a
lightning strike finds a short path to the ground or a stream in a mountainous
region finds the fastest path to the sea. Physicists have noticed these optimization
capabilities and have developed their own models of them - most notably
maximum entropy production principle. These thermodynamic models and
the Darwinian models cover similar territory and mutually illuminate each other.
Another area where Darwinian models have found to be applicable involves
observation selection effects. Spencer's survival of the fittest can be usefully
be generalized to
observation of the observable
- bringing observers into a
central location within Darwinism. Classical Darwinian models of selection
apply to observers in the exact same way that they apply to other entities.
One of the more studied areas of Darwinism within physics involves
quantum
physics. According to the many worlds interpretation, the world is constantly splitting.
Some branches reproduce faster than other ones, and so become more numerous and thus
more likely to be observed. Other interpretations propose a selection effect acting on these worlds
- known as "wave function collapse". Whichever interpretation is closer to the truth, this looks as
though this will be an interesting area for Darwinian or quasi-Darwinian models.
It has also been speculatively proposed that the visible universe is the product of Darwinian evolution - and thus
has a lineage of ancestors which existed before the big bang. If so, we might reasonably expect to find some clues relating to this evolutionary heritage. We can see that the visible universe had a birth date - but other evidence of an evolutionary history
currently remains elusive. We should do some further research on this possibility.
Darwinism is now over 150 years old. However, according to the picture here, the Darwinian revolution is only part way through. There's a lot of remaining revolution to go - and the process involves a lot more than just mopping up some creationists.
For references not hyperlinked to above, see the references here and here.
Parent and offsping often correlate with respect to their location.
It is possible to inherit a high-fitness location; one tree can inherit the
sunny side of the hill from another. But the significance of this inherited
variation is limited. A population can near-literally 'explore' a physical
space, if location is heritable and is linked with fitness. It may move along
gradients of environmental quality it may climb hills, or settle around water.
But to the extent that reproductive success is being determined by location per se
it is not being determined by the intrinsic features that individuals have. If
extrinsic features are most of what matters to realized fitness — if intrinsic
character is not very important - then other than this physical wandering,
not much can happen.
What can happen is that adaptations can develop. Lightning strikes can find the
shortest path to the ground, propagating cracks can locate weaknesses in materials
and drainage patterns can develop structures that efficiently drain basins.
The idea that concepts like 'fitness' and 'adaptation' apply to these kinds of
simple inorganic systems is a big deal for physics - and a big deal for Darwinism.
Of course in these kinds of system more than position is inherited. For
example, in electrical discharges, charge is also inherited. However position
is important - it is copied with high fidelity, it can often vary considerably
and many other properties can depend on it.
Godfrey-Smith attempts to draw a distinction between "intrinsic" and "extrinsic"
traits - and then claims that this affects the "Darwinian character" of processes.
However, traditional Darwinism has no use for such a distinction - all it cares about is
whether traits are inherited. If you look at axiomatic expressions of Darwinian
evolution, "intrinsic" and "extrinsic" don't get mentioned. That's because they
don't matter. They are irrelevant to most evolutionary theory. Heredity of traits
is what matters - not whether those traits are "intrinsic" or "extrinsic".
It's true that "intrinsic" traits can be more numerous than "extrinsic" ones.
However, that's no reason to single out "extrinsic" traits and exile them from
Darwinism. Darwinism makes no distinction between "intrinsic" and "extrinsic" traits.
The exact same rules about copying with variation and selection apply equally to both
types of trait.
Peter says "the significance of this inherited variation is limited". It seems to me that
the significance of this inherited variation is huge. It it wasn't for positional inheritance,
we would all have been born in the vacuum of space and died instantly. It may be
only "physical wandering" that means that we were born on the surface of a planet -
rather than in interstellar space - but it makes the difference between life and
death for all of us. Location is actually a very important property
that affects fitness. We should study how it evolves using good old-fashioned
evolutionary theory - it absolutely does apply.
This post is about Darwinian physics. In optimization theory, for evolutionary approaches to
work as effective search algorithm, the fitness landscape must not be too rough.
If fitness at nearby points in the fitness landscape are unrelated, an evolutionary
search will do no better than a random search.
In positional
inheritance the domain of fitness landscape maps corresponds to
a two or three dimensional physical space. Fitness usually depends
significantly on the location within that space.
It is pretty clear empirically that
fitness
at nearby locations is often correlated - i.e. fitness is heritable in cases of positional inheritance. However this leads to the question of: why is fitness heritable?
That question goes to the smoothness of nature. If you look at the world, nearby parts are often similar to each other. It isn't just neighbouring fitness that is similar - all neighbouring properties tend to be similar. Why is that the case? Here are three significant reasons:
Entropy increase causes many lumpy and rough phenomena to become more smooth and uniform.
Copying also causes like to associate with like. In a forest, the trees are similar because of genetic copying
processes.
Gravity causes air to associate with air, water to associate with water and rock to associate with rock.
It is a major force causing like to associate with like.
It's possible to argue that copying is the fundamental phenomena here -
and that gravity and entropy increases are the result of copying. Entropy
increase can also plausibly claim to be fundamental. Gravity cannot
claim to be fundamental.
I like the explanation in terms of entropy. Nature might not exactly abhor a gradient, but it isn't terribly keen on edges. There are a bunch of dissipative processes that gradually scuff, dissolve and erode them out of existence.
Here's a similar video of glass breaking in slow motion:
The video illustrates that cracks propagate from locations other than the branching tips. Some distance behind the tip of the crack is still a possible source of new (usually transverse) cracks. In a Darwinian model of splitting and recombining individuals, the entities that are evolving are crack tips - but this video illustrates that the notion of a crack tip has to be significantly extended in space if it is to result in a good quality model.
The resulting shatter pattern appears to be fairly heavily reticulated. It looks like a network - rather than a simple tree. However, appearances can be deceptive. If you look at the slow motion evolution, each crack forms from an existing crack - and there's a strict parent-offspring relationship that holds everywhere. In this case, the pattern of cracks forms a genuine family tree - something you might not guess at if looking at the resulting static fracture pattern.
Fitness landscapes are a common way of visualizing the relationship between variables associated with an organism
and fitness. The height of the landscape represents fitness and the domain over which the fitness landscape is defined
is composed of other variables associated with the organism. Most frequently the information associated with heritable
elements is used - but other variables affecting fitness could be included as well - such as environmental factors.
In this post, we will apply the idea of fitness landscapes to simple systems involving positional inheritance. Hopefully this will help to illustrate how the concept of 'fitness' applies to these kinds of system. To create a plot of fitness, we have to say what we mean by fitness. Fitness is a notoriously overloaded and slippery idea in biology - as was once explained in a book chapter titled "An Agony in Five Fits". Here, we won't proscribe any particular definition of fitness, but rather will show how to apply some common definitions of fitness.
The first aspect of measuring fitness is to define what entities you are measuring the fitness of. If there are multiple types of organism in a system, you have to say which one you are interested in tracking the fitness of. In simple systems involving positional inheritance this decision is often relatively simple: since there's only one main candidate entity. For example, with lightning strikes tracking the reproducing tips of the lightning are the obvious candidate. With stream systems, the branching tips of the streams themselves would be the most obvious object of study. With propagating cracks, the crack tips would be the object of study. With diffusion-limited aggregation, the available aggregation points would be what was tracked. In many of these cases, the precision of the available measuring instruments may be a factor in deciding exactly what entities are tracked.
Having selected the entities to be measured, the next thing to do is to decide how to measure fitness. Although there are many fitness metrics used for different purposes, we can categorize them in a few main ways. Fitness metrics can be:
Relative or absolute - depending on whether you are interested in relative success or absolute results;
Expected or actual - actual fitness measure growth rates while expected fitness can be calculated in advance;
Short or long term - the time horizon affects fitness measurements: offspring don't always result in grandchildren;
Generational or per unit of time - measuring growth in generational time units can sometime be useful.
These all apply to positional inheritance systems - though generational measures of fitness are not much use there. It's like using generational fitness metrics with bacteria. Bacteria are mortal and do have lifespans - but their lifespans are not much like the human three score years and ten. It's more a case that the bacteria live until they have a fatal accident - and the frequency of bacterial deaths are heavily determined by environmental factors. Thus, the lifespan of a bacterium is not usually a particularly useful or interesting figure. Using generational fitness with simple positional inheritance systems is a lot like this.
A fitness landscape is usually a plot of fitness over gene space. The peaks illustrate where well-adapted organisms are likely to be found. The roughness of the fitness landscape influences whether and how quickly evolving organisms will be able to find the peaks.
With simple positional inheritance systems, the 'genes' in question are positions - since position is one of the main things that is inherited in these systems. So the domain of the fitness landscape plot is usually simple two or three dimensional space. Fitness measures how likely branching or splitting is to take place at points in that space. Since reproduction typically requires resources, fitness can be reasonably expected to be correlated with resource availability.
It is common for fitness landscapes to change over time. As the environment changes, different genetic combinations are favored - and the fitness landscape shifts dynamically. With simple positional inheritance systems fitness landscapes tend to change in a predictable manner - the highest peaks tend to be systematically eroded. Because reproduction requires resources, takes place where resources are plentiful and depletes local resources, resource-rich areas will be systematically exploited and eliminated.
Fitness landscapes only track the parameters specified in their domain. If other factors affect fitness, the calculated fitnesses will not be accurate if these are omitted. For example, it is common to leave environmental factors out of fitness landscape plots.This can result in a lack of realism. With simple positional inheritance systems, fitness can also depend on more than positional factors. For example, consider a spreading fire. The reproduction rate of flames will be heavily influenced by positional factors - such as the local availability of combustible material. However other factors can also affect the rate of flame reproduction - such as the wind direction and the temperature - these are often a function of time as well as position. If there are more factors you can add them to the domain of the fitness landscape - but then you get a more complex plot in a higher dimensional space - which might not be so easy to make use of.
It's a commonplace observation that reproduction requires resources.
Reproduction often creates a transient, local resource depletion that decreases the chance of future reproduction in the short term. This "reproductive resource gap" is the topic of this article.
I've long known about this "resource gap". It seems to apply to many organic and cultural systems. Recently, I have been thinking about how general this principle is. It clearly doesn't apply to all creatures. For example a termite queen doesn't have a measurable reproductive gap - her reproductive system is a pretty continuous assembly line. However, for many creatures - from bacteria to elephants, there is a "reproductive gap" - where the mother has to accumulate resources after splitting and before splitting again.
Splitting typically produces offspring which are smaller than the mother. One hypothesis is that size is responsible for part of the effect - that bigger things are more likely to split than smaller things.
However, we know that - in many cases, the reverse relationship holds - i.e.: smaller things are more likely to split than bigger things are.
This is true for most organisms in the biosphere: there's a strong negative correlation between an organism's adult size and their reproductive rate. Smaller critters reproduce faster.
It's also true of very large objects. For planets and stars, smaller objects are more likely to break up than larger ones are - because the larger ones are held together more effectively by gravity.
Also, if erosion or corrosion are involved in the splitting - then these forces apply to the surface of objects - so again, small objects would be more strongly affected - because they have relatively larger surfaces.
Another perspective on the issue comes from considering a simple, common case. Some of the most common particle interactions in the universe consist of photons hitting dust particles. The photons form a a clear family tree with a few high energy photons near the root and enormous numbers of low-energy photons at the tips of the branches and a clearly-defined set of branching points - when the photons hit the dust particles and split.
In this system, the distance (or time) from one branching point to the next doesn't increase with proximity to the root. If anything, there's a tendency for collisions to be quickly followed by more collisions (if you just hit some dust there may be more things to hit nearby). This is another case where splitting tends to lead to more splitting - rather than to less splitting.
There are many other cases where splitting leads to more splitting. In a landslide, rocks that lave been stable for a long period of time might suddenly split many times in quick succession. There are many cases where splitting generates jagged edges and jagged edges result in more splitting. Beach pebbles are an example of this. Another similar case involves splitting reducing structural integrity. An egg is the stereotypical example, but similar considerations apply to many structures with membranes or skins. Breaching the outer wall leads to splitting and rapid disintegration.
At this stage a brief recap. A reproductive resource gap seems to be a pretty common feature of organic and cultural evolution - often the mother seems to need time to recharge. This applies to K-Selected creatures - and also to many r-selected ones - such as bacteria. However in many simple physical systems, there's often no reproductive resource gap. Instead, we find the opposite: splitting is likely to be followed by more splitting.
At this stage it might be tempting to conclude that the reproductive resource gap is an adaptation for managing limited resources - and that the reason simple systems don't have a resource gap is that they are degenerative systems which can't accumulate adaptations.
However, I've skipped over presenting some significant data. There are, in fact, simple physical systems that do exhibit a reproductive resource gap. One example involves raindrops in a condensation cloud. Smaller droplets have larger surface area to volume ratios - and so are more likely to be held together by surface tension. Another system involves falling ink droplets in water. The easiest way to explain this is with a video.
The video pretty clearly illustrates droplet reproduction - and a family tree of droplets.
However, there's a characteristic delay between one droplet reproduction event and the next. It is as though the falling droplets need to
build up some kinetic energy before they can reproduce again.
In my opinion, these examples demonstrate that the reproductive gap is more than just an adaptation.
There's a simple physics of needing to accumulate resources after splitting and before splitting again.
This is, I think, an interesting result in a poorly-studied area. Future investigations into the topic could look into how widespread this "reproductive resource gap" is in simple physical systems and what the interactions are between the simple physics of needing to accumulate resources before splitting and adaptations for resource management in organisms.
A largely unnoticed scientific revolution has occurred over the past forty-five years. The Darwinian paradigm has been successfully applied to numerous fields outside of biology: including the Social Sciences, the Behavioural Sciences and most recently to the Physical Sciences and even to Physics itself. This revolution promises an unprecedented conceptual unification of the many branches of science. Darwin Does Physics argues that the extension of the Darwinian paradigm has proceeded in parallel with a more widely recognized scientific revolution concerning the fundamental importance of information throughout science. Information is seen to enter nature in the form of ‘inferential systems’ which accumulate knowledge required for existence. Darwinian processes are identified as the physical implementation of inferential systems. The theoretical paradigms of physics have struggled over the past hundred years to accommodate the massive increase in scope of the known universe which Physics’ must explain. The Newtonian paradigm, which includes most of its modern theoretical structures such as string theory, has been found wanting. In an attempt to circumvent these problems some physicists are now developing theories in the Darwinian paradigm to explain the most basic processes of physics. These theories are explained and suggestions made for their development. If Darwinian theories are proved successful in explaining even the fundamental physical mysteries science will have achieved a wondrous unification of its many diverse branches.
John Campbell previously wrote the generally excellent book Universal Darwinism. Darwin Does Physics looks set to be a small step for John Campbell but a large leap for most of the rest of the evolution comminity. Let's hope that some people actually read it.
Hi. I'm Tim Tyler and this is a video about Darwinian physics.
In the 1980s there was a movement to incorporate elements from physics into evolutionary theory. It was claimed that not all of the complex-looking patterns of organisms were there in order to benefit the genes of their owners. Instead, some of them were the product of self-organizing systems. Reaction-diffusion patterns were invoked to explain zebra stripes and cheetah spots - challenging the idea that these were adaptations whose purpose was to improve camouflage. Cellular automata were found that reproduced the decorative patterns found on some seashells. The branching tree-like pattern that is found everywhere in biology was likened to dendritic frost patterns, electrical discharges and propagating cracks. The search was on for the new rules that governed these self-organizing systems.
Now, we have found many of these rules. A big and important one has turned out to be: Darwinian evolution via natural selection. So, now the time is ripe for Darwinism to claw back some of the explanatory territory that was lost to complexity theory - and it is time for it to perform some invasions of its own - into realms that have been traditionally occupied by physics and chemistry.
Darwinism is based on processes involving copying with variation and selection. It was once widely thought that copying of the required kind was unique to biological systems - and that before the origin of life copying fidelity was too poor to support any kind of Darwinian evolution - with any copying processes producing instead an error catastrophe and a mutational meltdown. This argument was made by A. G. Cairns-Smith in 1982 - for example. However, this idea has turned out to be dramatically wrong. It turns out that the high-fidelity copying suitable for supporting cumulative adaptive evolution is ubiquitous in simple physical systems.
Lightning strikes are one of the easiest examples of Darwinian physics to visualize. Lightning makes a branching tree. This is a family tree - with regions nearer to the trunk being the ancestors of regions nearer to the tips. If you ask what information is transmitted from parent to child in this case, the answer is that the offspring inherit their parent's position. They inherit this extremely precisely - down to the nearest millimeter.
Position isn't the only thing that is inherited in this way, other attributes - such as velocity and chemical composition - are inherited too, although this inheritance is often less precise.
Examples of Darwinism in simple physical systems are ubiquitous. Most tree-shaped structures are the product of copying with variation and selection. This includes many crystals, fractal drainage basins, propagating cracks and electrical discharges. Re-radiation of photons after hitting dust particles is one of the most common events in the universe and it follows the classical Darwinian algorithm - producing a family tree of photons with the older photons with higher energies being ancestral to their more numerous, but less energetic offspring.
Many other cases do not so obviously exhibit a simple tree. For example, raindrops divide, but they also join creating a network - rather than a classical phylogenetic tree. Of course there are also broadly similar examples of joining in classical evolutionary biology: gametes fuse together, parasites inject DNA into hosts, bacteria assimilate other bacteria and lineages join forces in symbosis. Evolution involves joins and mergers as well as splitting and subdivision.
In other cases, the phylogenetic tree is only visible if you take a historical perspective. For example, every asteroid has an associated ancestral tree of other asteroids, moons, planets and stars that it was formed from. However, you don't see the tree by looking at the asteroid. Only if you look back in time does the family tree become visible.
It's well known that organisms can be modeled as optimizing a fitness function. The same thing applies to lightning - it too is optimizing a function. It acts as a crack in space the seeks out the path of least resistance in its search for the ground. The fact that organic evolution acts as an optimization process is widely exploited by genetic algorithms. There, humans allocate a fitness function and let simulated evolution do their optimization work for them. Optimization features maximands - and these give evolution an apparent teleological character. Biologists can ask what wings are "for" - and then give a reasonable-sounding functional answer. Darwinian physics brings the concept of adaptation to the evolution of inorganic systems - and it becomes reasonable to ask what their features are "for".
Ultimately, both living organisms and lightning strikes can be modeled in the same optimization framework using the same maximand - by treating them both as maximizing entropy.
Entropy maximization is a formulation of optimization processes from within physics which some physicists are already familiar with. Some might ask what the advantages are of taking a Darwinian perspective over one based on entropy maximization. This is a complex question which risks going beyond the scope of this video, but briefly, entropy maximization and Darwinian evolution are largely equivalent ideas which make most of the same predictions - since nature's maximand is closely correlated with entropy production. These are mostly two different ways of looking at the same phenomenon. However, it is possible to select locally for other things apart from entropy maximization - including entropy minimization. Path dependency within a Darwinian framework provides a context which helps to explain these apparent deviations from maximum entropy production.
Lastly, Darwinism neatly explains observation selection effects. While these have long been considered to be part of physics, they are obviously a case of selection acting on observations. We can reformulate the concept of survival of the fittest as observation of the observable. This more general concept acts as a grand unified theory of selection - which neatly covers both Darwinian evolution in biology and observer selection effects in physics and many other areas of science.
Over the last two centuries, Darwinism has had a revolutionary impact on biology, leaving most of the theories that preceded it in the dust. Physics and chemistry are relatively new and unexplored frontiers for Darwinian explanations - but it seems clear that incorporating Darwinism into physics will involve some rewriting of textbooks, and a fair number of psychological paradigm shifts, as scientists gradually awaken to the idea.
Historically, Darwinism has had a hard time penetrating areas that go beyond inheritance of nucleic acids. For example, cultural evolution is still a topic mired in confusion and controversy. However, physicists are supposed to be intelligent people. Hopefully they will prove to be more open to Darwin's ideas than many social scientists have been - and less inclined to treat the Darwin enthusiasts as hostile invaders.
Observation selection includes selection between observers and selection within observers.
Selection within observers could be further subcategorized into unconscious filtering and deliberative filtering.
Unconscious filtering has dynamic and static components. Static unconscious filtering includes perceptual limitations.
Selection of observations within observers is rarely seen in the context of observation selection - and it seems to be a neglected topic. Sometimesobservation selection is used as a synonym of observer selection. This seems like bad terminology to me - it's important to distinguish between these different phenomena.
In universal Darwinism, many optimization processes in nature are seen as being Darwinian.
There are many examples of such optimization processes outside biology. Lightning finds the shortest path
between storm clouds and the ground. Cracks seek out the path of least resistance. Water on mountains
finds the steepest downhill path. Water on flat ground finds the path that maximally erodes the surface it flows over.
If you look at optimization techniques used in computer science, they include "genetic algorithms" and "memetic algorithms" - which are inspired by biology. However, there are also "gradient descent" and "simulated annealing" - which are named after relatively simple physical processes. These non-biological optimization techniques are simple and primitive - compared to biologically-inspired techniques - but they are still capable of performing useful work.
Physicists with knowledge of non-equilibrium thermodynamics typically use "maximum entropy thermodynamics" and the maximum entropy principle to explain these effects. The question physicists will probably have - when contemplating universal Darwinism - will be: what does it offer that maximum entropy ideas do not?
It is an excellent question, but one with some good answers, I think:
Universal Darwinism explains some cases where maximum entropy thermodynamics struggles. Increasing entropy isn't the only possible fitness function that can be applied to populations - and adaptations can locally favour other optimization targets in relevant selective environments. These are cases which universal Darwinism can handle but where maximum entropy thermodynamics doesn't typically help very much.
Lastly, maximum entropy thermodynamics and universal Darwinism are quite different-looking theories with different histories. Darwinism is better developed in many respects. It has better models of lock ins, sub-optimality, combining existing solutions and other phenomena. Physics can benefit from Darwinism's maturity.
Other types of optimization in physics
Another example of optimization in physics involves the opposite of hill-climbing: gradient descent. This can be demonstrated with a single ball bearing moving on a landscape - with little sign of a population, of copying, or of selective elimination.
Physics also features the principle of least action - which is probably its best-known optimization process. This works on the same principle as Galton's board (see right).
Putting a ball bearing into Galton's board computes the maximum value of the normal function - using gravity. The more iterative choices the ball faces, the better the board performs its optimizing task. The principle of least action is a broadly similar kind of optimization process. Galton's board illustrates iterated choices - but not terribly much reproduction. The balls and the gravitons involved are both produced at one location - and are clones of each other - but this copying seems rather remote from the optimization process itself.
An example of the principle of least action involves refraction of light. This case represents a challenge for population-level descriptions of optimization processes in physics. However, if considering a photon as a single point particle, refraction makes little sense. Only when a light wave is modeled as a spread-out, distributed phenomenon, can refraction can be understood.
The principle of least action hardly seems Darwinian at all. It can be modeled in simple physical systems devoid of much in the way of populations or copying. However, the search for shared principles that underlie the cases of optimization in physics may yet turn out to be a fruitful one.
The existence in mechanics of two actions and two corresponding variational principles which determine the true trajectories, with a Legendre transformation between them, is analogous to the situation in thermodynamics (Gray et al. 2004). There, as established by Gibbs, one introduces two free energies related by a Legendre transformation, i.e. the Helmholtz and Gibbs free energies, with each free energy satisfying a variational principle which determines the thermal equilibrium state of the system.
These other cases of optimization in physics raise the issue of what counts as optimization and what doesn't. Practically any physical system could be described as optimizing the function of behaving as it does - though such explanations may violate Occam's razor. The lightning strike in this post's illustration certainly looks as though it is performing search for high ground using a branching tree. However, we really need criteria relating to what qualifies and what doesn't.
Positional inheritance and velocity inheritance offer high fidelity transmission of information when physical objects split - allowing for adaptations to arise in physical systems. Copying with variation and selection in physics is a relatively simple and obvious application of Darwinism.
Darwinian physics is currently mostly an unknown topic in the mainstream. There's someQuantum Darwinism out there - but it is hard for most people to assess the worth of this material. Darwinian models apply well to fairly simple Newtonian systems. These models are more comprehensible and should be easier for most people to understand and accept.
Darwinian physics lacks some of the complications that cause social scientists to get into muddles about memes. For instance, simple physical systems have pretty undirected variation - allowing the standard neo-Darwinian assumption that mutations are random with respect to fitness to be applied - without much loss of modeling realism. The simple physical systems used as examples in Darwinian physics are easier to understand than the complex structures involved in human cultural evolution.
Darwinian physics has been worked on. Bickhard and Campbell's 2003 paper covers the topic - listing crystal growth, convection cells and catalysis as targets of explanation. D. B. Kelley's book on Universal Darwinism also covers it. However, for whatever reason, the subject area has not yet become mainstream knowledge.
Physicists are reasonable people. It seems reasonable to expect that they will regard the project of Darwinizing physics with less hostility than social scientists regard the project of Darwinizing culture.
Another reason to put energy into Darwinian physics is that physics has historically been a "high-status" science. If physicists come to accept that Universal Darwinism applies to basic inorganic physical systems, that will be a big and important feather in the cap of Darwinism.
Social science is still the area of greatest social and political importance for evolutionary theory. It's the area where there's the greatest need for correct, Darwinian science. It's the area where backwards, pre-Darwinian science is doing the most damage. It's an area where religious attempts to distort science get a lot of funding. However, Darwinian physics can contribute to this project indirectly - by helping to apply increased pressure to the social scientists on multiple fronts.
I previously nominated
positional
inheritance as the most common kind of inheritance.
The second-most common kind is probably velocity inheritance. The term
"Velocity inheritance" refers to the way in which entities that split tend to inherit their parent's velocity. The offspring pieces tend to inherit the velocity of their parents.
Turbulence illustrates velocity inheritance fairly clearly. Or think of the way the targets break up in clay pigeon shooting.
Velocity inheritance is quite a lot like positional inheritance - but there are some differences:
Positional inheritance is always high-fidelity. However, the fidelity of velocity inheritance breaks down considerably when the particles involved become very small. In particular if the offspring particles are neutrinos or photons, velocity inheritance is not very high fidelity. You can also see poor velocity inheritance if you smash small objects into much larger ones at enormous speeds.
Some may find that velocity inheritance is easier to understand than positional inheritance. Velocity is more obviously a property of objects that is copied when they divide. Others find it more confusing - and get caught up in (irrelevant) concerns about relativity and frames of reference.
Other "simple" properties are inherited too when entities split - mass, angular momentum, charge, etc. The inheritance is not not always which such high fidelity as is seen in position or velocity inheritance.
Hi. I'm Tim Tyler, and this is a video about incorporating observers and observations into Darwinism.
Universal Darwinism generalizes many of the principles of evolutionary theory, giving it a broader range of applicability and extending it to cover the inanimate world. One such principle is natural selection. Universal selection applies broadly to the natural world, harvesting asteroids and atoms - as well as ants and aardvarks.
A closely-related concept that can be similarly generalized is "survival of the fittest". Richard Dawkins, writing on page 12 of "The Selfish Gene", wrote:
Darwin's survival of the fittest is really a specific case of a more general law of survival of the stable. The universe is populated by stable things.
This proposed law of "survival of the stable" has subsequently not received proper attention. However it too is a specific case of a more general law, which might be referred to as: "observation of the observable".
Dealing with observability implicitly includes all cases of survival - since, in order to be observed, entities must have survived. However it expands the scope of original idea in two ways - to include:
Potential observables being hidden, and...
Observers failing to observe fully;
With this modification, Darwinism can be accurately applied to entities which are merely hidden - instead of ones that have failed to survive. This allows evolutionary theory to be applied to cases such as the Tall poppy syndrome - where entites are selected on the basis of their visibility - rather than because their competitors failed to survive.
Survival - from "survival of the fittest" is an important determinant of what we observe, but it is not the only factor involved.
The new principle also incorporates selection effects acting on observers and their perceptual apparatus into evolutionary theory. Such effects have classically been studied in the context of "the anthropic principle". However, the term "anthropic" was always unfortunate - since it implies the effect only applies to humans - whereas, in fact, any observer will do: animals and machines can act as observers too. Universal Darwinism brings observation effects under the umbrella of evolutionary theory. It includes cases where observers are sleepy, deluded, lacking awareness, or have perceptual limits.
Darwinism is the most appropriate framework for modeling the effect of selection on observations and observers. The expansion of Darwinism to deal with "observation of the observable" is long overdue. Previous iterations of the theory have made no mention of observations or observers, but this development puts them back into Darwinism at its heart. To fully explain scientific observations, we have no choice but to use the more general and broadly-applicable principle.
Hi. I'm Tim Tyler, and this is a video about positional inheritance.
In universal Darwinism, copying is found ubiquitously in nature, from spreading ripples to propagating cracks, from growing crystals to scattering radiation. Copying - in conjunction with variation and selection - forms the basis of Darwinian evolutionary theory.
The copying in universal Darwinism includes DNA, culture, and a whole bunch of other aspects of the environment. To give some examples of environmental inheritance, rabbits inherit rabbit warrens, corals inherit their coral reef - and so on. The most common form of environmental inheritance is positional inheritance. To give some examples of this:
Raindrops - split and produce offspring that inherit their parents' position.
Cracks - have dividing tips and offspring crack tips start their lives near to their parents.
Atoms - split during nuclear decay - and the offspring particles originate near the parent atom.
Because of the property of physics known as locality, any form of inheritance is also accompanied by positional inheritance. That makes positional inheritance the most widespread form of inheritance in existence.
The products of positional inheritance often form tree-like structures. The roots and branches of plants resemble trees - and actually are phylogenetic trees of plant cells, laid down in order during development - in a combination of phylogeny and ontogeny. Similarly, lightning, propagating cracks, fractal drainage patterns, and crystalline dendrites are all associated with prominent visual trees. In each case, these are family trees, that show the path of descent. Sometimes the associated phylogenetic trees are less obvious. For example, in a landslide, each moving boulder has been pushed into motion by collisions with one or more parent boulders. Though each boulder can trace its ancestry back to the first falling stone, the resulting family tree is not obvious to casual observers. It's the same with raindrops in clouds and vortices in turbulent fluid flow.
Positional inheritance also results in adaptation - another hallmark of Darwinian evolution. Cracks adaptively seek the weakest path through matter, streams adaptively trace out the boundaries of their associated drainage basins and turbulence selectively forms where there is the most energy to feed it.
One thing that evolving systems typically need, in order to exhibit complex adaptations, is high-fidelity copying. Excessive noise often results in inherited information getting lost - and this leads to the disintegration of complex adaptations. However, positional inheritance often has pretty high fidelity - allowing complex adaptations based on it to remain stable.
Though positional inheritance is a pretty central concept in Darwinian evolution, it is a curiously neglected idea. While some seem to appreciate that organisms inherit their parents' environment as well as their genes, simple inheritance of position gets practically no recognition as a component of evolutionary theory. Sad times for Darwinism.
 We now at last have a significant academic literature on meme-gene coevolution. However few seem to have considered the dynamics of the meme-gene-queme coevolution that can be expected as a result of considering quantum Darwinism in the brain. This blog post is a brief attempt to share my thoughts on the topic.
We now at last have a significant academic literature on meme-gene coevolution. However few seem to have considered the dynamics of the meme-gene-queme coevolution that can be expected as a result of considering quantum Darwinism in the brain. This blog post is a brief attempt to share my thoughts on the topic.
 I came up with the idea of branch tip evolution in 2012 by thinking about
I came up with the idea of branch tip evolution in 2012 by thinking about 




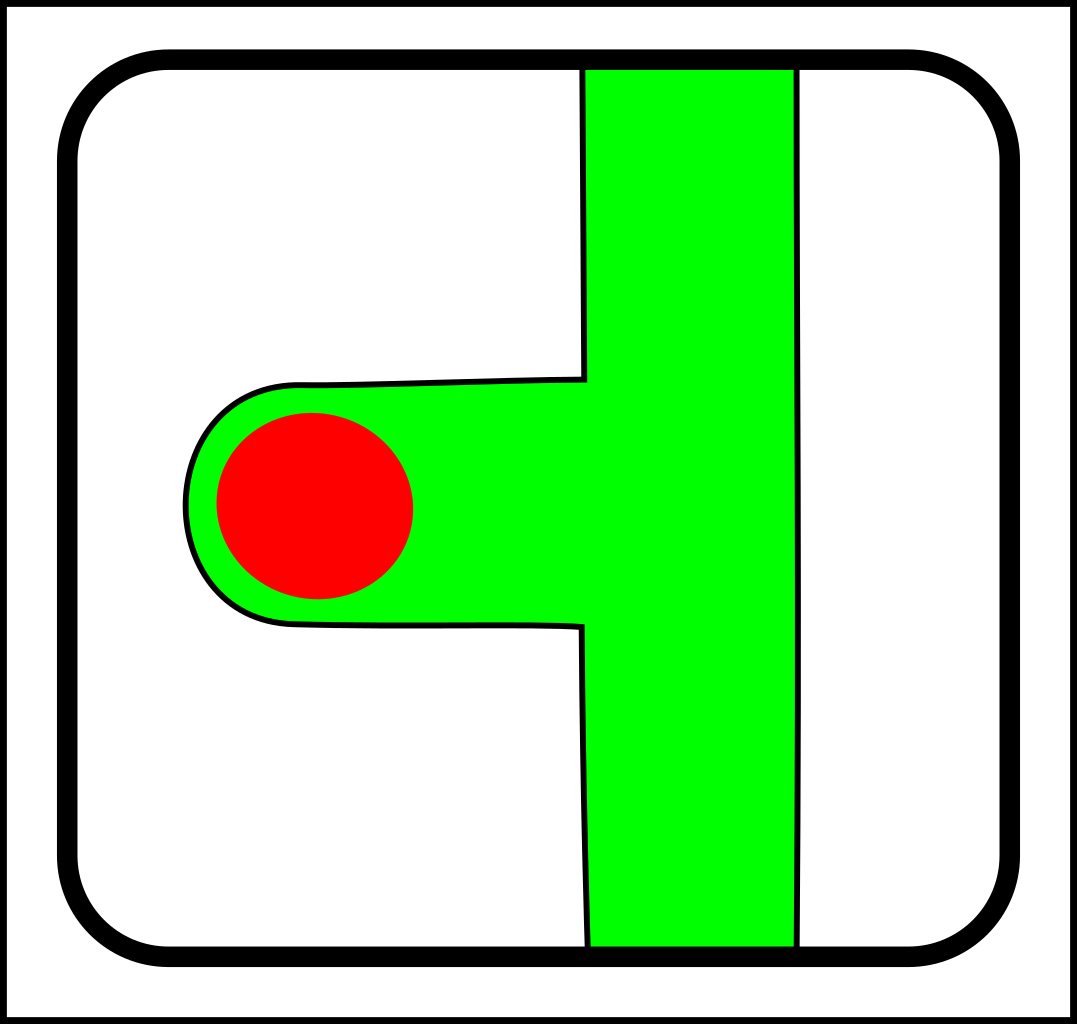



 One of the key concepts in understanding these systems is
One of the key concepts in understanding these systems is
 I notice that Peter Godfrey-Smith has a section in
I notice that Peter Godfrey-Smith has a section in  This post is about Darwinian physics. In optimization theory, for evolutionary approaches to
work as effective search algorithm, the fitness landscape must not be too rough.
If fitness at nearby points in the fitness landscape are unrelated, an evolutionary
search will do no better than a random search.
This post is about Darwinian physics. In optimization theory, for evolutionary approaches to
work as effective search algorithm, the fitness landscape must not be too rough.
If fitness at nearby points in the fitness landscape are unrelated, an evolutionary
search will do no better than a random search. Fitness landscapes are a common way of visualizing the relationship between variables associated with an organism
and fitness. The height of the landscape represents fitness and the domain over which the fitness landscape is defined
is composed of other variables associated with the organism. Most frequently the information associated with heritable
elements is used - but other variables affecting fitness could be included as well - such as environmental factors.
Fitness landscapes are a common way of visualizing the relationship between variables associated with an organism
and fitness. The height of the landscape represents fitness and the domain over which the fitness landscape is defined
is composed of other variables associated with the organism. Most frequently the information associated with heritable
elements is used - but other variables affecting fitness could be included as well - such as environmental factors.

.gif) In
In  Universal Darwinism explains some cases where maximum entropy thermodynamics struggles. Increasing entropy isn't the only possible fitness function that can be applied to populations - and adaptations can locally favour other optimization targets in relevant selective environments. These are cases which universal Darwinism can handle but where maximum entropy thermodynamics doesn't typically help very much.
Universal Darwinism explains some cases where maximum entropy thermodynamics struggles. Increasing entropy isn't the only possible fitness function that can be applied to populations - and adaptations can locally favour other optimization targets in relevant selective environments. These are cases which universal Darwinism can handle but where maximum entropy thermodynamics doesn't typically help very much.
 I previously nominated
I previously nominated














