In computer science, filtering and sorting are big topics. Donald Knuth devoted volume 3 of his epic The Art Of Computer Programming to these two topics. That's a reasonable indication of their importance in computer science.
However, science in general took a different route. Filtering is dealt with partly as "selection" - which is covered most comprehensively by evolutionary biology. However filtering is a much broader topic, which extends well beyond biology. As a result, the science of selection is fragmented:
Anthropic reasoning - a form of observation selection.
Because the science involved is fragmented there are also a number of areas where it could be applied, but currently isn't - because its influence is not understood or recognised. When small coins accumuate in your wallet, that's a type of selection. Similarly selection results in unpalatable goods accumulating in your refridgerator. Selection is also important in many common physical phenomena, such as erosion, crack propagation, catalysis, crystal growth and electrical discharges. However, its influence often goes unrecognised there as well.
In 1971, George Price called for a theory of selection, writing:
A model that unifies all types of selection (chemical, sociological, genetical,
and every other kind of selection) may open the way to develop a general
‘Mathematical Theory of Selection’ analogous to communication theory.
Price continues with:
Selection has been studied mainly in genetics, but of course there is much
more to selection than just genetical selection. In psychology, for example,
trial-and-error learning is simply learning by selection. In chemistry, selection
operates in a recrystallisation under equilibrium conditions, with impure
and irregular crystals dissolving and pure, well-formed crystals growing. In
palaeontology and archaeology, selection especially favours stones, pottery, and
teeth, and greatly increases the frequency of mandibles among the bones of
the hominid skeleton. In linguistics, selection unceasingly shapes and reshapes
phonetics, grammar, and vocabulary. In history we see political selection in
the rise of Macedonia, Rome, and Muscovy. Similarly, economic selection in
private enterprise systems causes the rise and fall of firms and products. And
science itself is shaped in part by selection, with experimental tests and other
criteria selecting among rival hypotheses.
If the situation with filtering in science is bad, the situation with sorting is surely worse. At least selection is championed by evolutionary biologists. Sorting is also very common. You can see its results while looking at stones on a beach or clouds in the sky. Shaking your breakfast cerial makes the biggest lumps rise to the top - a simple sorting operation. I have talked about "natural sorting " before - but most people have never heard of it. If the science of filtering is fragmented, the science of sorting is positively obscure.
There have been efforts to build a general science of selection. Proponents of universal Darwinism have been working on it. There's Zukav's "Without Miracles". There's Hull's "Science and selection". There's Fog's "Towards a universal theory of competition and selection". There's Campbell's "Epistemological roles for selection theory". It is probably fair to say that most of the pieces are out there, but the topic is far from penetrating the scientific mainstream. It seems as though more work in the area remains to be done.
Positional inheritance has become one of my key concepts. It's important when explaining universal Darwinism, because it is common, simple and easy to understand and visualize. However, my previous writings on the topic have been spread out over many web pages. Here are the main points, collected from my previous writings and organized. The page is low in hyperlinks and animations - since this is a draft of a book chapter on the topic. However, pictures still illustrate the main points.
Positional inheritance
We have previously seen that copying is found ubiquitously in nature, from spreading ripples to propagating cracks, from growing crystals to scattering radiation. Of course, copying, variation and selection are the basis of Darwinian evolutionary theory. The copying can take a variety of forms, but the most basic is positional inheritance.
It is common knowledge that people inherit the environment of their parents - along with their parents genes. They inherit the local climate, the local language, government and religion - along with traits coded in DNA. A number of parental traits are inherited in these examples, but one of the attributes which is always inherited is position.
It is common for organisms to inherit their parents' position with considerable precision. Not all organisms have effective dispersal strategies - so often the apple does not fall far from the tree. Rabbits tend to inherit the warren of their parents. Corals inherit their parent's reef - and so on. Much the same is true of many inorganic natural forms.
Examples
Here are some examples of inorganic positional inheritance:
Raindrops - split and produce offspring that inherit their parent's position.
Cracks - when a crack tip divides the offspring crack tips start their lives nearby.
Atoms - when atoms split, the offspring particles originate near to the parent atom.
Ripples - parent ripples give rise to child ripples near to their parents.
Because of locality in physics, any form of inheritance is also accompanied by positional inheritance. That makes positional inheritance the most widespread form of inheritance in existence. Positional inheritance applies to waves and particles of all kinds. Since waves and particles are important building blocks of the universe, this makes positional inheritance very widespread.
Trees
The products of positional inheritance often form tree-like structures. The roots and branches of plants resemble trees - and actually are phylogenetic trees of plant cells, laid down in order during development - in a combination of phylogeny and ontogeny. Similarly, lightning, propagating cracks, fractal drainage patterns, and crystalline dendrites are all associated with prominent visual trees. In each case, these are family trees, that show the path of descent. That these trees are in fact family trees can often be easily verified by filming their formation in slow motion. Videos of lightning strikes slowed down show that forks always descend from existing branches. If you look at videos of bullets hitting panes of glass you will see that propagating cracks behave in a similar manner - the cracks spread outwards in a radial pattern where each crack descends from an earlier parent crack.
Sometimes the associated phylogenetic trees are less obvious. For example, in a landslide, each moving boulder has been pushed into motion by collisions with one or more parent boulders. Though each boulder can trace its ancestry back to the first falling stone, the resulting family tree is not obvious to casual observers. It's the same with splitting raindrops, vortices and photons. Family trees are still involved, but you would need a time lapse image to see them.
Heritable fitness
One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, relatively fit offspring should be ancestral to relatively fit descendants. Without this condition being met, adaptations can't get off the ground. Does positional inheritance exhibit heritable fitness? Often, fitness is heritable in systems involving positional inheritance - simply as a result of the uneven distribution of resources needed to fuel division.
For example, in diffusion-limited aggregation systems, the concentration of aggregating particles is often greater in some places than others. In electrical discharge systems, the potential gradients can be greater in some places than others. With propagating cracks, the medium can be more brittle in some places than others. These situations are all commonplace ones. In each case, the association between fit ancestors and fit descendants is due to what might be called the smoothness of nature: the tendency of natural systems to be locally fairly uniform on a small scale - the tendency for nearby places to be alike.
In short, ancestors who are in the right place at the right time tend to have descendants who are in the who are in the right place at the right time - while
ancestors who are in the wrong place at the wrong time tend to have descendants who are in the who are in the wrong place at the wrong time. The reason is simple: the descendants are born near to the ancestors and so tend to share a similar environment. The result is heritable fitness.
Fidelity
One thing that evolving systems typically need, in order to exhibit complex adaptations, is high-fidelity copying. Excessive noise often results in inherited information getting lost - and this leads to the disintegration of complex adaptations. However, positional inheritance often has pretty high fidelity - allowing adaptations based on it to remain stable. If you think of it in terms of coordinates in the universe that are unknown to an observer, by learning the location of an object an observer gains a considerable quantity of information - that object's coordinates in three dimensional space. If the offspring is within 1 meter of the parent, then that's about 265 bits of mutual information copied with high-fidelity. Say 350 bits of spacetime. Of course in practice, few positional inheritance systems take up the whole universe, so 350 bits is an upper limit. 350 bits is peanuts compared to biological systems, but it represents a search space of considerable size - it's enough for some non-trivial optimization processes to take place.
Adaptation
Positional inheritance also results in adaptation - another hallmark of Darwinian evolution. Cracks adaptively seek the weakest path through matter, streams adaptively trace out the boundaries of their associated drainage basins and turbulence selectively forms where there is the most energy to feed it.
Some cases which are easy to understand can be found in the organic realm. A tree growing partly underneath a bridge is a useful example. Where the branches are under the bridge they don't grow so well, due to lack of light. The parts of the tree that are not under the bridge grow more vigorously. The result is an adaptive fit between the tree and the bridge. This adaptation is not caused by to changes in DNA. It can happen even if every cell in the tree is genetically identical. With a tree under a bridge the adaptive fit between the tree and its environment is caused by differential reproductive success of the cells of the tree - their different rates of growth, reproduction and death. However the important evolving variable is not stored chemically the tree's cells - rather it is the position of the cells themselves which affects their fitness.
If it is acknowledged that the goodness of fit between the tree and the bridge is an adaptation, then by the same logic we ought to count a number of inorganic systems as exhibiting adaptations too - since they display essentially the same dynamics. Crystal dendrites growing near a heat source adapts to grow around the hot area. Rivers and streams adapt to avoid rocky outcrops, cracks propagate around reinforced areas - and so on.
Multiple inheritance channels
Since the modern evolutionary synthesis traditional evolutionary theory has specialized in studying the evolution of nucleic acid-based creatures. A splinter group has rebelled against this orthodoxy, promoting dual inheritance theory - the idea that there are two main information highways in biology - one which transmits inherited information via cells and the other which transmits inherited information down the generations using brains and social learning. However, two inheritance channels is just not enough. Information can also be transmitted down the generations using multiple other channels. Velocity, time, chemical composition and electrical charge are among the many other variables that can be inherited.
Temporal inheritance merits a mention here. Positional inheritance only covers the three dimensions of space. However since Einstein's era, science has understood that space and time are interwoven, and that it often makes sense to talk about spacetime. Spatio-temporal inheritance is a bit of a mouthful, though. Also, it makes reasonable practical sense to talk about positional inheritance and temporal inheritance separately.
Including temporal inheritance and velocity inheritance - which are both also common - bumps up the information carrying capacity of many simple physical systems, making their evolutionary dynamics more interesting.
Universal inheritance
Positional inheritance makes inheritance ubiquitous in the universe. Far from being confined to biology, inheritance happens whenever starlight hits dust - one of the most common interactions in the universe. This is part of the justification for using the term "Universal" in "Universal Darwinism". A number of other writers on the topic have expanded Darwinism to culture, but left the theory confined to biology - leaving chemistry and physics out of the domain of Darwinism. Others have embraced Darwinism in physics - but only applied it to quantum theory, observation selection or the possibility that our entire visible universe might have ancestor universes that existed before the big bang. These applications of Darwinism are interesting, but still narrow, making "Universal Darwinism" not very "universal". Here, Darwinian evolutionary theory is expanded to most dissipative structures - making its application domain much larger and making the theory correspondingly more significant.
Limitations
Positional inheritance doesn't lead to adaptations on the scale seen in biological evolution. Various problems and limitations reduce its scope for generating adaptations. Many inorganic systems exhibiting positional inheritance lack important properties found in the organic domain.
One such property is an unlimited number of generations. Living organisms today can trace their lineage back four billion years. By contrast, many positional inheritance systems last for tens, hundreds or thousands of generations before going extinct. Electrical discharges or propagating cracks and splitting photons are all examples of positional inheritance systems which tend to have definite origins and finite lifespans. I some cases, the finite lifespan is a product of the lack of a growth phase. In biology, organisms typically divide and then grow before dividing again. Not all positional inheritance systems have this growth phase. Some divide, divide and divide again. Rocks are usually like this and so are photons. By contrast electrical discharges and drainage basins do have a growth phase. Without a growth phase, unlimited inheritance is obviously impossible. Even with a growth phase, inorganic systems often have a limited temporal extent. Lightning strikes feature a growth phase, but it is fueled by a finite potential difference, and once the potential gradient diminishes, the lightning's pathway disappears.
Another limitation involves the quantity of material inherited. Living organisms can transmit megabytes of information to their descendants. However positional inheritance just doesn't support such large quantities of information. 10 - 50 bits seems like a more reasonable figure for many positional inheritance systems. You can still communicate using 10-50 bits - but the bandwidth limit restricts what can be said.
Criticism
Peter Godfrey-Smith has a section in Darwinian Populations and
Natural Selection denigrating the significance of positional inheritance.
He writes (on page 55):
Parent and offspring often correlate with respect to their location.
It is possible to inherit a high-fitness location; one tree can inherit the
sunny side of the hill from another. But the significance of this inherited
variation is limited. A population can near-literally 'explore' a physical
space, if location is heritable and is linked with fitness. It may move along
gradients of environmental quality it may climb hills, or settle around water.
But to the extent that reproductive success is being determined by location per se
it is not being determined by the intrinsic features that individuals have. If
extrinsic features are most of what matters to realized fitness — if intrinsic
character is not very important - then other than this physical wandering,
not much can happen.
What can happen is that adaptations can develop. Lightning strikes can find the
shortest path to the ground, propagating cracks can locate weaknesses in materials
and drainage patterns can develop structures that efficiently drain basins.
The idea that concepts like 'fitness' and 'adaptation' apply to these kinds of
simple inorganic systems is a big deal for physics - and a big deal for Darwinism.
Godfrey-Smith attempts to draw a distinction between "intrinsic" and "extrinsic"
traits - and then claims that this distinction affects the "Darwinian character"
of processes - with extrinsic traits not being very "Darwinian". However, most
traditional evolutionary theory has no use for such a distinction - all it cares
about is whether traits are inherited. If you look at axiomatic expressions of Darwinian
evolution, "intrinsic" and "extrinsic" inheritance don't get mentioned. That's because
this is a distinction that doesn't make much difference: it is irrelevant to most
evolutionary theory. Inheritance of traits is what matters - not whether those traits
are inherited via "intrinsic" or "extrinsic" mechanisms.
Peter says "the significance of this inherited variation is limited". It seems to me that
the significance of this inherited variation is huge. It it wasn't for positional inheritance,
we would all have been born in the vacuum of space and died instantly. It may be
only "physical wandering" that means that we were born on the surface of a planet -
rather than in interstellar space - but it makes the difference between life and
death for all of us. Location is actually a very important property
that affects fitness. Evolutionary theory is mostly agnostic about how information
is passed down the generations - so we can use its existing tools to study positional
inheritance.
Here's a draft of a book chapter I have written on the topic of observer and observation evolution.
Introduction
One of the places where mainstream physics has come closest to embracing Darwinism involves the role of
observers. Physicists identified the role selection of observers plays, and identified it as the cause of goodness
of fit between man and his environment - in the form of life-friendly physical laws and a stable planetary home.
History
Historically, the evolution of observers was first studied by a physicist called Brandon Carter
in the 1970s. One of the ideas he came up was was that physical constants having values that
made the universe habitable was not due to chance or an intelligent designer. It was no accident
that we observed a universe with life-friendly physical laws, since it could not be otherwise:
any observers in universes with physical laws that were not life friendly would rapidly perish.
This idea was christened "the anthropic principle" and it was contrasted with the Copernican
principle - which states that we are not in a special place in the universe. According to the
anthropic principle we are in a very unusual place in the universe - one suitable for the
evolution of humans over billions of years.
The result of the need for life-friendly physical laws is an adaptive fit between the
universe and living systems inside it. The physicists didn't describe what they had
discovered as an "adaptive" fit. Instead they said that the universe was 'fine tuned'
for life. Most of the physicists involved didn't seem to link these ideas to evolutionary theory.
Instead they seemed to consider it to be an entirely new area of science which they had discovered,
one that could explain the appearance of design without invoking a designer. They did, however
use the terminology of 'selection' to describe their findings - mirroring the terminology used in
evolutionary theory. Retrospectively, it seems obvious that they were just applying Darwin's
discovery to human observers.
Observation selection
A subsequent development was the discovery that a similar idea could be applied to observations,
as well as to observers. Observations may be filtered in a number of ways - both before and after arriving
at the senses. For example, publication bias filters information before it reaches an observer. An
observer's expectations and preconceptions might then go on to filter information further before
it reaches consciousness.
Observation of the observable
From one perspective, observation selection is one type of selection in the nervous system among
many. Filters also control whether information is stored, when it is retrieved, and
when it is forgotten. Still more filters are applied to ideas, action plans and motor outputs. However,
if you look at the situation another way, observations seem to be fundamental - since all
knowledge gains are made through observation. Observations are the basis of everything an organism
knows. Everything else consists of inferences derived from observations. This gives observations a
primary status. For example, if a fruit fly in an scientists's experiment dies, that's a case of natural
selection. However the scientist doesn't learn about it until an observation is made. We can say that
survival of the fittest is a special case of observation of the observable. This results in a reformulation
of evolutionary theory puts observers at the heart of the theory. This broadly mirrors the changes in
physics that happened at the turn of the last century, when it was discovered that observers played a
surprisingly central role in physics.
Observation reproduction
Selection is only part of Darwinism. That many observers reproduce is perhaps too obvious to mention.
However, observation reproduction merits a few comments. Behavioral reproduction is ubiquitous in
human cultural transmission. It is widely agreed that humans often copy the behavior of other humans
via behavioral imitation. However, behavior is only one side of behavioral imitation. To be copied, behaviors
have to also be observed. Observations reproduce during this process just as behaviors do. To give an
example of an observation that catalyzes its own reproduction consider the observation of yourself,
snorting cocaine. Such an observation is often followed by more similar observations. This is a simple
case of observation reproduction.
Beyond survival and reproduction
Observation selection illustrates how Darwinian dynamics can involve more than survival and reproduction.
Observation selection also filters out things that are hidden from the observer. This allows evolutionary theory
to be applied to cases where the observation of entities is based on their visibility - rather than because they
and their ancestors survived and reproduce. Survival and reproduction are important determinants of what
we observe, but they are not the only factors involved.
Multiverse
The idea that the visible universe is the product of selection effects suggests that the
visible universe is part of a multiverse. It which case it would be helpful to know the size
of the multiverse - and which parameters are free to vary in it. Alas we can only observe our small
corner of the multiverse. This leads to a difficult inductive inference problem with very little
data to go on. The concept of a "reference class" is sometimes used to denote the set of objects
being selected from. When dealing with other possible worlds it isn't always clear what the
set of worlds being selected from consists of - since we only see one world.
Brandon Carter's Ultra-Darwinism
One of the physicists who did recognize links to Darwinism was the originator the idea in the first
place: Brandon Carter. He wrote in 1992 that: "anthropic selection should be considered as an adjunct
to ordinary natural selection". He proposed that the union of anthropic selection and natural selection
be called 'Ultra Darwinism'. That's pretty much the same thing that I am saying - except that Brandon
used a different name. However, I would emphasize that the topic is mostly just applying basic
Darwinian principles to observers and observations. There are a few other topics involved too -
for example, the maximum entropy principle is used to handle ignorance. However, this is mostly
ordinary Darwinism applied to observers and observations.
Terminology
The term "anthropic" turns out to be rather unfortunate. The "anthro-" prefix means:
"man", but the basic idea can easily be generalized to cover animals, plants and machines. The
human-based version of the idea seems anthropocentric to the point of being unscientific.
Then again, look at the rock called granite. It is composed mostly of grains of feldspars and quartz, with some mica and other minerals inserted among them. When granite weathers, the feldspars and micas become clays but nothing much happens to the quartz grains. They are most resistant and get transported down streams or along shores. Thus most beaches are the result of differentially eroded granite. This is an example of natural selection in the nonliving world. Quartz grains survive longer than feldspar grains, and there is a progressive increase in the average resistance to weathering, of the set of grains that have still survived. This action of natural selection is even creative as we see by the formation of a beach. The lack of reproduction imposes constraints on the flexibility of evolution here, but one shouldn't confuse that with the selection itself. We do have here a common sort of evolution by natural selection and there are many other non-living examples.
Of course, the case for Darwinan physics is quite a bit stronger than this passage implies - because copying and reproduction are also common in the inorganic realm. Rocks split into smaller rocks, streams split into smaller streams, and so on. There's also evidence of family trees - as seen in diffusion limited aggregation, and optimization and exploring a search space - as when a lightning strike finds the highest point in a landscape. However, Van Valen had some of the important ideas quite early on in the history of the field.
The term "refactoring" commonly refers to a set of useful computer programming techniques which involve making structural changes to a program without altering its function. Refactoring is often done before making functional changes. If you launch into making changes without some preliminary refactoring, you are often more likely to make mistakes. Refactoring frequently allows you to make changes without breaking tests - so you can verify that your changes don't break anything. If you need to make a bunch of changes, it sometimes helps to make ones that you can verify do no damage first.
I argued in refactoring science that science can benefit from refactoring too. I agree with the many folk who think that Darwinism is in need of some changes. This raises a number of issues about what changes need making. One such issue is where to start from. Darwinism has had two main identifiable releases: Darwin's Darwinism and the Modern Synthesis. Many seem to assume that we should build on the most recent release of the theory, the Modern Synthesis. For example, here's Massimo Pigliucci:
It makes just as little sense to talk of "Darwinism" in modern science as it does to talk about Mendelism or Newtonianism. The current theory of biological evolution is the Modern Synthesis, and if one wants to make the point that cultural evolution works in the same way, one needs to take on board the most refined version available of the theory, not its earliest draft.
Incidentally, I think that's why we should avoid talk of Lamarckism as well as Darwinism altogether: they refer to murky (in the first instance) or outdated (in the second) ways of thinking about biology, and it doesn't help to resurrect them as if the last two centuries of science hadn't happened.
This rather assumes that the Modern Synthesis was an improvement over Darwinism. However, some have argued that the Modern Synthesis consists of a bunch of overstatements. For example, Mesoudi (2011) has argued that cultural evolution is Darwinian, but not Neo-Darwinian. The modern synthesis is often billed as uniting Darwin's idea of natural selection and Mendelian genetics. The problem is that Mendelian genetics only applies to inheritance via DNA. As a result, other forms of inheritance were sidelined by the synthesis. From such a perspective, the Modern Synthesis looks like a bit of a broken version of Darwinism. Failing to encompass cultural evolution is a pretty serious flaw. In this case, maybe we should start from an earlier revision of Darwinism - one without so many problems.
Starting with Darwin's theory should allow cherry picking any useful bits from the Modern Synthesis. Starting from the Modern Synthesis would probably be followed by bunch of 'revert' operations - to get rid of the dogmatic and mistaken bits. The former operation looks easier to me than the latter one.
I have some other concerns about starting from the Modern Synthesis. It is vague. Of course Darwin changed his mind about some things too - and the various editions of "The Origin" describe rather different theories. However, it seems to me that Darwinism is more clearly specified than the Modern Synthesis was. The Modern Synthesis can be difficult for critics to criticize - since there's no canonical version. In science, that's not a good thing.
In some respects, the Modern Synthesis illustrates how not to produce an evolutionary synthesis. It subsequently became a bit of a straitjacket for researchers. Inevitably any new theory of evolution will also be incomplete - but there's no need for it to be dogmatic and mistaken.
Darwinism's domain has expanded dramatically over the last century, to cover culture, development and other fields. This has led many thinkers to wonder how far the Darwinism goes. Universal Darwinism proposes that it goes far. This page offers a brief introduction to the basic ideas involved.
Basics
First, some basics: evolutionary theory is based on population thinking. It considers the world as composed of a population, an environment, and operations that act on the population members. It assumes that part of the world can be broken down into a set of similar entities - a population. A finite set of simple operations can then be applied to these entities. Entities can come into existence, go out of existence, make copies of themselves and merge with other entities. They can self-modify, interact with one another and with their environment. They can be filtered and sorted. They can move around and can be moved around. Meta information associated with each operation explains where, when and under what circumstances it is to be applied. The operations are usually applied repeatedly over an extended period of time.
Coevolution
In practice sometimes it is important to consider more than one population. Where multiple populations of entities interact, models of coevolution and symbiosis can be applied. These consist of descriptions of the additional populations, together with more operations describing how the entities from different populations interact.
Simplified models
Many models are simplified and focus on a small set of these operations, often ignoring other ones completely. For example, they may assume that interactions take place randomly, that the population is of infinite in size, or that the environment is fixed. Sometimes the simplifications assist mathematical analysis, other times they make computer simulation easier. Darwinism is not a particular theory, rather it is a toolbox which can be used to construct theories. One thing that is out of bounds is "arbitrary" mutations. Unconstrained mutation predicts everything and is useless. Mutations must be constrained to be naturalistically plausible. Other operations are subject to similar constraints.
Expanded domain
Traditionally, the populations involved consist of living organisms with genes made out of nucleic acid. The organisms involved are usually of the same species. Universal Darwinism expands the domain of evolutionary theory to include a wide range of other types of populations. Often these are conventionally considered to be part of the environment. For example, in culture, populations of cars, books, buildings, coins and words can be considered. In neuroscience, populations can consist of nerve impulses, or axon branch tips. In geology, populations can consist of rocks, streams, mountains or islands. In physics, populations can consist of atoms, molecules, photons or observers.
Often the same kinds of operations that are conventionally associated with Darwinism can be applied in these new domains. Operations involving damage and disintegration clearly apply to a very wide range of entities. Also, many entities persist and can be copied. Copying is sometimes though of as being unique to living organisms, but it isn't so: copying is ubiquitous in nature. The essence of copying is that information in one place winds up in more than one place - and that happens whenever rocks crumble, whenever streams part and whenever starlight hits dust. High fidelity copying is also common. For instance, position is often copied with high precision when entities split.
As a result of these shared operations, many of the parts of evolutionary theory can be applied to gain insights into these other realms. In particular, the concepts of fitness, adaptation and optimality which are traditionally associated with evolutionary biology turn out to apply widely to many types of system.This is Universal Darwinism in a nutshell.
Is the approach useful?
How useful is evolutionary theory in these other domains? It varies. In the case of cultural evolution, the answer is obviously, very useful indeed. In other cases, it often depends partly on how much overlap there is between the operations involved and those found in classical Darwinism. One common issue is limited heredity. With both DNA and culture, high fidelity inheritance systems allow open ended evolution and the accumulation of adaptations. However, with many simple systems, heredity is limited. A rock might inherit its position, momentum, chemical composition and temperature from its parent rock, but there's a limit on the number of times a rock can split into smaller rocks. Additionally, there are ceilings on the information carrying capacity of some entities. The tip of a lightning strike might carry information about charge and position - and encoding position might require a considerable number of bits - but there's still a limit on the amount of information so encoded. Such "information ceilings" represent additional limitations. These limitations don't mean that many standard Darwinian methods are not useful - but they should lead to constrained expectations regarding the scale of any resulting adaptations.
Relevance of population thinking
Another issue is the relevance of population thinking in the first place. With DNA-based organisms, a population of discrete similar entities is often the product of evolution - since the entities involved share a common ancestor. Often they flock together and interact with each other more than with other parts of their environment. In some of the other cases covered by Universal Darwinism, how to divide the world into entities is not always so clear and obvious. Even if the entities involved don't form a natural kind, it can still be useful to divide the world up into pieces and consider how they interact - that is simply standard scientific reductionism. However, the more the world divides neatly into pieces, the easier it will be to apply evolutionary theory to it. Note that even if you can't divide some aspect of the world up into pieces, you can still sometimes gain something by treat it as a single persisting entity - and then applying coevolution models.
Competing theories
A theory is only as good as the best of its competition. How have the phenomena covered by universal Darwinism been treated by scientists historically? We will consider two alternative theoretical frameworks:
One competing theory is from physics, or more specifically thermodynamics and statistical mechanics. This is the maximum entropy production principle. Instead of considering fitness maximization, maximum entropy production deals instead with entropy maximization. Maximum entropy production covers similar domain to universal Darwinism and successfully reproduces many of its predictions. However there are differences in emphasis and approach. For example, evolutionary theory has done a lot of useful work on path dependence and genetic drift. It has also made more study of when waste arising from conflict results in deviations from optimality.
Another competitor for universal Darwinism is Niche Construction Theory. Like universal Darwinism this deals with heredity transmitted via the environment and selection acting on environmental components. Niche Construction Theory is usually presented as a bunch of add-on components for Darwinism, though. Universal Darwinism produces very similar results by using standard Darwinism with no more axioms or complexity. Instead it treats the environment as a bunch of other Darwinian populations and reuses models of how different populations interact. As such, it would seem to be favored by Occam's razor. If Universal Darwinism is adopted first on these grounds, it is not obvious that there is any further explanatory work for Niche Construction Theory to do.
Deepening as well as broadening
Many biologists have long hoped that we will discover alien life - to illuminate which aspects of biology are fundamental and which are historical accidents. Universal Darwinism provides a very similar kind of illumination. In addition to the broadening
of the domain of Darwinism, the new examples of evolution stimulate deepenings, revisions and generalizations of evolutionary theory. For example, cultural evolution stimulates the incorporation of intelligent design and directed mutations into evolutionary theory. I haven't covered these sorts of deepenings in this post - mainly through shortage of space - but several such deepenings are conceptually associated with Universal Darwinism.
Darwinism: what's in a name?
Some debate whether the resulting evolutionary theories should be called "Darwinian". Sometimes certain operations - such as entity copying - are required for the term to be applicable. Other times certain operations - such as inheritance of acquired traits - are forbidden. The term is used here mostly as a way of paying homage to Darwin's pioneering work on organic and cultural evolution. The term Darwinism is commonly used, though it has some associated baggage. In the end this is a terminology issue - the science involved is more significant.
How revolutionary?
Another issue is how revolutionary Universal Darwinism is. On one hand, most evolutionary textbooks need to be completely rewritten to cover the new domains involved. On the other hand Universal Darwinism expands the domain of Darwinism at very little cost in terms of additional axioms or complexity. It is often no more complex and traditional Darwinism. From this perspective it is no revolution at all. Having said that, expansions of the domain of Darwinism often represent revolutions - in the fields which are being invaded. For example, Darwinism represents significant revolutions in anthropology, economics and physics. In biology, the field was significantly transformed after adopting Darwinism. It seems reasonable to expect a similar large impact in other fields.
I came up with the idea of branch tip evolution in 2012 by thinking about positional inheritance. Probably the simplest way to understand it is by thinking about the evolution of buds on tree branches. Buds reproduce and they undergo selection. The result is often an adaptive fit between the shape of the tree and its environment. For example, some buds may be in shade under a bridge while others may overhang a busy road. Selection affects the growth and reproduction of the buds, resulting in an adaptive fit between the shape of the tree and its environment.
Copying with selection resulting in adaptive fitness are the hallmarks of Darwinism. However this was not a form of Darwinism based on DNA genes. The example holds up even if all the tree buds involved are precisely identical in terms of their DNA.
A kind of Darwinism of branch tips can be useful in explaining a wide range of tree-shaped structures in nature. In the organic realm, there are branches, roots, corals, circulatory systems, respiratory systems, and branching axons and dentrites. Inorganic tree-shaped systems include electrical discharges, propagating cracks, crystal growth, and drainage basins.
Many models of these types of inorganic system take a functional approach to them - for example, saying that drainage basins form efficient structures for rapidly removing water from landscapes - of that they maximize the production of entropy. From an evolutionary perspective, such models are all very well, but they are all to do with adaptive function. Adaptation is part of evolutionary theory - but it also has another side: path dependence, or historical contingency. Evolutionary theory provides a rationale for adopting a functional perspective in the first place, and it also helps to explain cases where there are deviations from what strict functionalism might predict.
While branch tip evolution is an excellent and important model of many physical systems, it has limitations. In real trees, there is cell reproduction within the branches as well as at the branch tips. Also, there are other cases where branch dynamics are important - i.e. when not all the action in the system is taking place at the branch tips. Finally, real branch tips can sometimes shrink as well as grow. In such cases, the analogy between the real tree and a family tree starts to break down. More sophisticated models involving graph evolution may be a better fit for such cases.
Through the first half of the 20th century evolution was mostly the science of selection and splitting. To illustrate with diagrams, a simple selection operation looks like this:
->
->
...and splitting looks like this:
->
->
Then in the 1960s, symbiosis and sex started to look as though they needed another fundamental operation - merging - which looks like this:
->
->
These are the basic operations in modern evolutionary theory. As you can see from the diagrams, these basic operations are particulate - in that they operate on discrete, particle-like entities.
I'm thinking that a node-and-link scheme would be a better match for the evolution of brains, tree branches, circulatory systems, immune systems, respiratory systems - and so on.
At first glance, this might seem like a case of adding operations that create and destroy links between nodes. That might make sense in computer science, but in nature, links form gradually and a more gradualistic scheme seems more in tune with existing evolutionary theory. Here are the splitting operations:
->
->
->
->
->
->
...and here are the corresponding joining operations:
->
->
->
->
->
->
For the sake of simplicity, I've only shown diagrams for nodes with fewer than four links. I've also
omitted selection events - i.e cases where elements (nodes or links) are simply destroyed.
I should probably say a bit more about the motivation for such a scheme. In biology, tree and graph evolution
is often reducible to particle evolution - since the components involved are often composed of particulate cells.
However, this doesn't work so well for brain evolution - since axons and dentrites are so link-like.
It's often possible to model branch tip evolution using conventional Darwinian models which treat branch tips as
individuals. This works quite well in many of the types of inanimate systems considered by Darwinian physics, for example. However this approach works best when flow into the resulting tree-like structures is time-like - i.e. unidirectional and at a constant speed. If resources can flow back out of unpromising roots or newly-shaded branches, conventional Darwinian models based on a population of branch tips do not work so well. Then, more sophisticated models can be useful.
There is a field called evolutionary graph theory -
but that seems to be about something rather different from the topic of this page.
I just noticed that Addy Pross offered a criticism of universal Darwinism in 2011:
we routinely attempt to explain psychological phenomena in biological terms, biological phenomena in physical and chemical terms, chemical phenomena in physical terms, and so on, not the other way around. The observation of Darwinian-like behavior at the chemical level is highly significant, not because it suggests that molecules behave in a biological fashion, but because it opens up the possibility of explaining biological behavior in chemical terms
Addy goes on to say:
As mentioned above, the temptation to interpret the behavior of molecular replicators in biological terms - fitness, natural selection, survival of the fittest, etc., should be firmly resisted. Chemical phenomena are more usefully explained in chemical terms
My response would be that this line of argument is based on a misidentificaton of where evolutionary theory lies in the scientific pecking order. It is true that evolutionary theory is used in biology - but it is more properly identified with statistics - in particular with statistical mechanics. This puts it above physics - along with systems theory, chaos theory, information theory, statistical thermodynamics and other theories that transcend physical law.
Addy's proposal that we should start with chemistry is a complete non-starter - since any chemical explanation would rather obviously fail to explain how Darwinism applies to physics and computer science.
Tree-shaped patterns in nature are frequently found in both the organic and inorganic domains and
demand a common explanation. One type of explanation is a functional one. Many trees
efficiently drain basins. Electrical discharges can be modeled as draining electrical
charge and propagating cracks do something similar with stress. The maximum entropy production
principle generalizes this idea so that it can be applied to evolutionary family trees
as well as to physical trees.
An alternative approach to modeling natural tree-shaped systems is to enumerate the type of operation
that are possible on them, and develop specific theories for each type of operation.
This is the approach taken by Darwinism - which breaks the dynamics of family trees
into reproduction with selection, mutation and recombination. Darwinism also applies to
branch tip dynamics in other tree systems - since branch tips split, mutate,
undergo selection and inherit properties from their parent(s). At the very least they
inherit their position.
I've argued elsewhere that positional inheritance is ubiquitous and that this means that many inorganic systems exhibit Darwinian dynamics. For example, the tree-shape propagating cracks, electrical discharges and fractal drainage basins are family trees. In these systems, the branching tips of the trees form a population which is subject to copying with variation and selection. As a result conventional Darwinian concepts of fitness and adaptation apply - and these systems act as optimizers in a similar way to what is seen with genetic algorithms.
With positional inheritance, most physical tree-shaped patterns in nature are regarded as being family trees. However the fact that these trees are physical trees (rather than being historical ones) means that their branches might undergo dynamic changes. Because a family tree is historical its branches form a fixed structure which can't be changed once it has formed. However, the branches of other natural tree-shaped patterns (e.g. streams or with electrical discharges) sometimes do change their structure over time. The branches don't just grow at the tips, they wave and break.
What this means is that we need a theory of branch dynamics - in addition to the
Darwinian dynamics associated with the population of branch tips.
Basic branch operations that do not change branch topology include:
Branch widening;
Branch narrowing;
Branch lengthening;
Branch shortening;
In addition, branches can be destroyed and sometimes entire branches may also split off or join together. Branches can break without becoming detached if the tree is reticulated (as is fairly common in natural systems).
Streams illustrate branch widening, narrowing, lengthening and shortening. Streams get wider as more rain falls and they narrow in times of drought. In meandering rivers on plains, streams lengthen. Branch separation is illustrated by crystals and coral. Sometimes a chunk breaks off without being destroyed. It might go on to start a new colony elsewhere. Branch death sometimes follows branch separation. It can also be a consequence of extended branch shortening or narrowing.
Specific theories relating to these branch operations are generally domain specific. However there may be commonalities between the operations in different domains. Modeling such commonalities is part of the study of branch dynamics, though it is beyond the scope of this post.
Tree dynamics are of theoretical interest because trees are common natural objects. It is also noteworthy that it is possible to recover traditional Darwinism from the theory by considering the special case where the tree branches become fixed once they are created and only the branch tips evolve. The principles of tree dynamics are a superset of those in Darwinism.
Various folk have expressed concern that a
generalized version of Darwinism that covers more
phenomena would be weaker and would constrain expectations
less - and would therefore be less useful.
John Maynard Smith once wrote:
The explanatory power of evolutionary theory
rests largely on three assumptions: that mutation is
non-adaptive, that acquired characters are not inherited,
and that inheritance is Mendelian - that is, it is atomic,
and we inherit the atoms, or genes, equally from our two
parents, and from no one else. In the cultural analogy,
none of these things is true. This must severely limit
the ability of a theory of cultural inheritance to say
what can happen and, more importantly, what cannot happen.
I am not convinced by the way-too-easy claims of “everything evolves!”,
such as here, mainly because it seems to me that broad conceptions of
evolution tend to have less explicative power
I've previously replied to Maynard Smith, writing:
Let's assume for a moment that his conclusion is true -
and that it is harder to make predictions with cultural evolution
than it is with biological evolution.
So what? Theories of cultural evolution are not in competition with
theories of biological evolution - they compete with other theories
of cultural change that are less inspired by Darwinism.
Here I want to offer a different response - which is nonetheless still
based on the idea that theories that cover different domains are not
direct competitors.
Darwinism can be axiomatized. The axioms can be weakened and/or discarded,
resulting in cut-down versions of Darwinism that apply under different
circumstances. For example, conventionally, Darwinism requires copying.
However without copying you can still have selection and goodness of fit -
which are important components of Darwinism. Another common constraint
involves what counts as a mutation. Random mutations constrain expectations
a lot and result in easily falsifiable theories.
However under some circumstances, such a constraint on mutation is
not realistic. In cultural evolution, for example, it can be quite
reasonable to model mental mutations as consisting of practically
any change (short of recombination) that can happen inside an
individual's mind. Mutations are still relatively small - but
since they are the result of multiple generations of copying
and selection within an individual's mind, they can be adaptive.
Other Darwinian axioms can also be usefully weakened.
However, the resulting broad theories aren't in direct competition with the
earlier narrow ones. They are applicable in different domains. As a result
we have an ensemble of more-or-less Darwinian evolutionary theories -
which are applicable under different circumstances. Having these
extra theories in no way weakens the old, narrow versions of Darwinism
from the textbooks. Those still work just as they did before. However the
new, broader theories extend evolution into new realms, such as physics,
chemistry, geology, astronomy and computer science.
Here's a diagram of the ensemble of evolutionary theories:
Theories vary along a one-dimension axis from 'specific' to 'general'.
The more general theories are vaguer and constrain expectations
less. However their corresponding advantage is that they are
still applicable in application domains where narrower theories fail completely.
The expanded domain of evolutionary theory represented by
universal
Darwinism is like having a bunch of new tools in your toolbox.
The complaints of John Maynard Smith and Alberto Acerbi are
a bit like complaints that some of the new tools aren't as useful
as one of the old tools was. That's OK - these are new tools that
we didn't have before. You don't have to throw out any of your old
tools in order to make room for the new ones. Instead, take delight
in all the new things the bigger toolbox lets you do.
Of course this doesn't address the terminological debate about what
deserves to be described as being 'evolutionary' or 'Darwinian'.
However that's more of a 'small peanuts' debate, in my humble opinion.
Universal
Darwinism gives us a bunch of shiny new tools. We should try them
out, see what they can do, and learn when best to use them.
The domain of Darwinian evolution has expanded dramatically over the last 150 years.
Darwinian evolutionary theory is now frequently applied to cultural evolution, the
development of individuals and individual learning. More speculative extensions of
Darwinism include ones that cover quantum physics, complex adaptive systems,
cosmological natural selection and observation selection effects.
It is natural for observers of this expansion to ask: how far can the Darwinism
go? Where is the the edge of evolutionary theory? What are the limits of Darwinism?
The questions relating to "Darwinism" might be criticized as being a bit vague - but we
can replace these with similar questions about the domain of concepts such as fitness
selection and adaptation - and have some more rigorous questions that more people are
likely to be able to form a consensus about.
I think that the lesson of history is that the edge of evolution is farther out than
we think. People are inclined to say that the edge of Darwinism lies at the outer
edge of their personal understanding of it. However, we can see historically that
Darwinism has repeatedly pushed into new domains, covering new phenomena.
Another possible position is that there's no real "edge" - instead evolutionary
theory gradually breaks down as more and of its axioms are progressively broken.
I think that it is clear that there's some truth to this perspective. However,
evolutionary theory is fairly simple - and there aren't very many axioms to
break. Nonetheless, we should not necessarily expect to find a single
precipice at the edge of evolution - but rather a gradual disintegration
in the form of some steps or a slope. This complicates the issue - but
doesn't fundamentally alter the problem.
To finish this article, I have a characterization of where I think the edge
of evolution is to offer. I think evolutionary theory applies to macroscopically
irreversible systems. This gives it roughly the same domain as
maximum entropy thermodynamics - which
I claim
it is very similar to. Part of the intuition behind this involves the link between selection
(from evolutionary theory) and destruction (which leads to many macroscopic entropy increases).
This relationship is probably wrong in detail. There's nothing in evolutionary
theory that forbids its application to macroscopic reversible systems. Selection
need not necessarily be linked to destruction. However, this is the best, short
characterization of the edge of evolution that I have. Without it, I am reduced
to offering a laundry list of phenomena that I think that evolutionary theory applies to.
It's well known that Donald Campbell was an early pioneer of cultural evolution. Most in the field acknowledge his work as influential or historically important. One of his early papers on "Blind variation and selective retention" dates from 1960. Later in life, Donald Campbell explored the limits of evolutionary theory - applying it to a range of inorganic phenomena. In particular, there's the following paper:
Here, Campbell and Bickhard apply the principles of variation and selection to a range of phenomena - including why
gravel accumulates at the edges of roads, crystal growth, crystal stability, the formation
of atoms and molecules and catalysis. They argue that "energy wells" are common causes of selection phenomena - giving rocks, planets, stars, and galaxies as examples.
Campbell mostly avoids the terminological debate about whether such phenomena qualify as being "Darwinian" - which is still the source of much modern noise and confusion. Instead he uses terminology oriented around the terms "variation" and "selection" and
examines whether they produce "goodness of fit", "adaptation" and "evolutionary historicity".
There seem to be some differences between my understanding of this area and Campbell's (which admittedly dates from the 1990s). I would give examples of tree-like phenomena in nature - such as electrical discharges, propagating cracks, fractal drainage patterns and diffusion-limited aggregation - and say they these trees are family trees, showing clear ancestor-descendant relationships. I make that argument in more detail on my positional inheritance page.
Another apparent oddity of Campbell's perspective is that none of his examples of selection seem to involve things being destroyed. Death is a common source of selection in the organic realm - so it seems natural to me to give instances of destruction as examples of selection in the inorganic realm. For example, the destruction of islands by the sea, of rocks by landslides, of sediments by subduction, and of pebbles by erosion all seem to me to be fine examples of selection. Campbell seems to me to systematically steer clear of destructive examples. However, this seems like a very curious thing to do. Did Campbell really not regard destruction as being a source of selection?
Though Campbell still seems to me to have a bit of a restricted perspective on the scope of selective explanations, he applied the concepts of selection, fitness and adaptation deeply into the realms of physics and chemistry in the 1990s. That makes him into one of the pioneers of Universal Darwinism.
Here's a similar video of glass breaking in slow motion:
The video illustrates that cracks propagate from locations other than the branching tips. Some distance behind the tip of the crack is still a possible source of new (usually transverse) cracks. In a Darwinian model of splitting and recombining individuals, the entities that are evolving are crack tips - but this video illustrates that the notion of a crack tip has to be significantly extended in space if it is to result in a good quality model.
The resulting shatter pattern appears to be fairly heavily reticulated. It looks like a network - rather than a simple tree. However, appearances can be deceptive. If you look at the slow motion evolution, each crack forms from an existing crack - and there's a strict parent-offspring relationship that holds everywhere. In this case, the pattern of cracks forms a genuine family tree - something you might not guess at if looking at the resulting static fracture pattern.
Replace repetitive expressions by calls to a common function
This is a type of operation known as "refactoring". Refactoring - for any non-programmers in the audience - involves reorganising code without changing its function. I think what's currently happening with memetics is a similar type of operation involving refactoring science.
A very common type of refactoring operation involves identifying two pieces of code that perform similar functions and replacing them with calls to a common subroutine. Separate pieces of code that perform similar tasks can arise in many ways. Similar code could be developed independently by different developers. Or it could be duplicated from a shared source and modified for a new purpose.
In science we see essentially the same thing: models are developed independently, turn out to have essentially the same dynamics and then need combining.
A classic recent example of this involves kin selection and group selection. While originally conceived as very different processes, many modern formulations have turned out to be different ways of expressing the same types of dynamics. Group selection and kin selection have turned out to be close synonyms.
Organic evolution and cultural evolution are currently getting the same treatment - in that "universal" models are being developed that cover both cases.
Part of the motivation for this type of refactoring is normally that it prevents duplicated maintenance work. When maintenance effort needs duplicating, it costs more to perform. The branches involved can gradually get further out of step with each other as time passes. This introduces incompatibilities and merging the branches can become increasingly expensive as time passes.
As with this type of refactoring in computer science, the original duplicated routines do not need to be performing exactly the same function as one another. Even if they are doing a similar job it often pays to combine them. Sometimes the differences are represented as different parameters. Sometimes they are "lambda functions". Sometimes the differing functionality is encapsulated in pluggable modules.
That's the role that genetics and memetics play in evolutionary theory. They are pluggable modules that are accepted as parameters to a more general evolutionary theory.
That it clearly proposes this refactoring operation is one of the unique features of memetics. It neatly partitions the required changes when adapting evolutionary theory to cover culture into:
Changes to evolutionary theory it make it more general;
The creation of encapsulated theories of genetics and memetics;
I think that some of the debates over memetics are illuminated by this comparison with refactoring - at least for those
with a background in computer science. When refactoring, there are often team members that say features should be being worked on instead. Sometimes the objection that refactoring will introduce bugs is made. Others point to the cost of the refactoring operation. Some say that the code isn't that similar after all, and shouldn't be combined. Some say it's too late to make the change at this stage, and we should learn to live with the old design.
I think we see many of the same objections being made by those involved in evolving modern evolutionary theory. However, this does seem like a pretty attractive refactoring to me. It is worth bearing in mind that science is forever. We should strive to make our models clean and beautiful - for the sake of those that come after us.
Why does this commonality between computer programming and science exist? I think that's a fairly easy one: both science and computer programs involve building and maintaining models of the world - and that's enough to explain the commonality.
Selection has been studied mainly in genetics but of course there is much more to selection than just genetical selection. In psychology for example trial-and-error learning is simply learning by selection. In chemistry, selection operates in a recrystallisation under equilibrium conditions, with impure and irregular crystals dissolving and pure, well-formed crystals growing. In paleontology and archaeology selection especially favours stones, pottery, and teeth and greatly increases the frequency of mandibles among the bones of hominid skeletons. In linguistics selection unceasingly shapes and reshapes phonetics grammar and vocabulary. In history we see political selection in the rise of Macedonia Rome and Muscovy. Similarly economic selection in private enterprise systems causes the rise and fall of firms and products. And science itself is shaped in part by selection with experimental tests and other criteria selecting among rival hypotheses.
This is from Price's posthumously-published 1995 manuscript, "The Nature of Selection". Price went on to lament the lack of a proper theory of selection and the lack of a proper definition of selection. He compared the current state of selection theory with the state of information theory in 1922. Price died in 1975 and apparently this paper was written in 1971 - which makes it ahead of its time indeed. Extensive quotes from "The Nature of Selection" are available free online - in the paper George Price's Contributions to Evolutionary Genetics.
While things have moved on a bit since 1970s the theory of selection is still little known. Evolutionary theory should cover it - but hardly anyone seems to realize that the theory applies to physical systems like crystals in the way that Price clearly indicated. Evolutionary theory is still in an immature state, where it isn't applied through much of its rightful domain. Progress is pretty treacle like: Darwinism has a pretty huge inertia. Also there are fragmented areas of science that are part of the study of selection that are scattered all over the place. Observation selection seems to be studied mostly by physicists and links to evolutionary theory are mostly ignored. Selection bias and sampling effects are concepts which are widely used in science - but again, these topics are rarely seen as being in the domain of evolutionary theory.
It's a commonplace observation that reproduction requires resources.
Reproduction often creates a transient, local resource depletion that decreases the chance of future reproduction in the short term. This "reproductive resource gap" is the topic of this article.
I've long known about this "resource gap". It seems to apply to many organic and cultural systems. Recently, I have been thinking about how general this principle is. It clearly doesn't apply to all creatures. For example a termite queen doesn't have a measurable reproductive gap - her reproductive system is a pretty continuous assembly line. However, for many creatures - from bacteria to elephants, there is a "reproductive gap" - where the mother has to accumulate resources after splitting and before splitting again.
Splitting typically produces offspring which are smaller than the mother. One hypothesis is that size is responsible for part of the effect - that bigger things are more likely to split than smaller things.
However, we know that - in many cases, the reverse relationship holds - i.e.: smaller things are more likely to split than bigger things are.
This is true for most organisms in the biosphere: there's a strong negative correlation between an organism's adult size and their reproductive rate. Smaller critters reproduce faster.
It's also true of very large objects. For planets and stars, smaller objects are more likely to break up than larger ones are - because the larger ones are held together more effectively by gravity.
Also, if erosion or corrosion are involved in the splitting - then these forces apply to the surface of objects - so again, small objects would be more strongly affected - because they have relatively larger surfaces.
Another perspective on the issue comes from considering a simple, common case. Some of the most common particle interactions in the universe consist of photons hitting dust particles. The photons form a a clear family tree with a few high energy photons near the root and enormous numbers of low-energy photons at the tips of the branches and a clearly-defined set of branching points - when the photons hit the dust particles and split.
In this system, the distance (or time) from one branching point to the next doesn't increase with proximity to the root. If anything, there's a tendency for collisions to be quickly followed by more collisions (if you just hit some dust there may be more things to hit nearby). This is another case where splitting tends to lead to more splitting - rather than to less splitting.
There are many other cases where splitting leads to more splitting. In a landslide, rocks that lave been stable for a long period of time might suddenly split many times in quick succession. There are many cases where splitting generates jagged edges and jagged edges result in more splitting. Beach pebbles are an example of this. Another similar case involves splitting reducing structural integrity. An egg is the stereotypical example, but similar considerations apply to many structures with membranes or skins. Breaching the outer wall leads to splitting and rapid disintegration.
At this stage a brief recap. A reproductive resource gap seems to be a pretty common feature of organic and cultural evolution - often the mother seems to need time to recharge. This applies to K-Selected creatures - and also to many r-selected ones - such as bacteria. However in many simple physical systems, there's often no reproductive resource gap. Instead, we find the opposite: splitting is likely to be followed by more splitting.
At this stage it might be tempting to conclude that the reproductive resource gap is an adaptation for managing limited resources - and that the reason simple systems don't have a resource gap is that they are degenerative systems which can't accumulate adaptations.
However, I've skipped over presenting some significant data. There are, in fact, simple physical systems that do exhibit a reproductive resource gap. One example involves raindrops in a condensation cloud. Smaller droplets have larger surface area to volume ratios - and so are more likely to be held together by surface tension. Another system involves falling ink droplets in water. The easiest way to explain this is with a video.
The video pretty clearly illustrates droplet reproduction - and a family tree of droplets.
However, there's a characteristic delay between one droplet reproduction event and the next. It is as though the falling droplets need to
build up some kinetic energy before they can reproduce again.
In my opinion, these examples demonstrate that the reproductive gap is more than just an adaptation.
There's a simple physics of needing to accumulate resources after splitting and before splitting again.
This is, I think, an interesting result in a poorly-studied area. Future investigations into the topic could look into how widespread this "reproductive resource gap" is in simple physical systems and what the interactions are between the simple physics of needing to accumulate resources before splitting and adaptations for resource management in organisms.
One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, fit offspring should be ancestral to fit descendants. Without this condition being met, adaptations can't get off the ground.
In this post, I will argue that this requirement is commonly met by many types of simple, natural systems
involving positional inheritance.
The thesis here will be that macroscopic variations in resources are common - and result fairly directly
in heritable fitness. If resources are very evenly distributed, then the condition that fitness is heritable would not be met.
Such extremely even distribution of resources can happen if the environment is near to equilibrium, for example.
With diffusion-limited aggregation systems, the concentration of aggregating particles can be greater in some
places than others. In electrical discharge systems, the potential gradients can be greater in some places
than others. With propagating cracks, the medium can be more brittle in some places than others. These situations are
all commonplace ones.
The existence of heritable fitness is consistent with observed adaptations in these types of systems. Drainage basins
are well adapted to rapidly dissipate the potential energy in the incoming rainwater - and form similar structures to
drainage systems designed by engineers. Lightning strikes take the shortest path from the cloud to the ground. Cracks
seek out lines of weakness - resulting in an adaptive fit between the actual cracks and the weak points of the material.
In practice, the requirement for heritable fitness is a pretty trivial condition which is almost always met.
To evolve adaptations some additional, more stringent conditions are also required. Essentially, the selection pressure
needs to out-weigh the mutation pressure. If it doesn't do so, you get an error catastrophe - and no adaptations.
In other words, devolution - rather than adaptive evolution.
One criticism of Universal Darwinism is that the core principles it is based on - namely heredity, mutation and selection - are too general, are capable of explaining anything and therefore explain nothing.
Neo-Darwinism avoids this issue - by claiming that mutations are "undirected". However, this is not really an acceptable approach in cultural evolution - where it is widely agreed that mutations are not remotely random - and that modeling them as being "undirected" is a coarse and limited approach.
I generally attribute the correct resolution of this issue to Donald Campbell. He basically said that variation was generated on the basis of existing knowledge. He used the unfortunate term "blind" to describe this - which has led to endless misunderstandings of his idea by confused critics.
Gary Cziko's terminology from the title of his book, without miracles is another approach to the problem. The term "without miracles" expresses the idea that mutations should be explicable naturalistically. While naturalistic explanations are good, scientists normally take it for granted that their explanations should be plausible within a naturalistic framework. The idea that mutations do not involve "miracles" isn't really all that much of a constraint - and the more scientific theories constrain expectations the more valuable they are.
This article tries to answer the question of what the minimum constraints on mutation are that avoid the accusation that the resulting theory is vacuous. The answer is pretty simple: all that is necessary is that mutations should be limited. If an evolutionary sequence requires mutations that exceed the mutation limit, they violate the predictions made by the theory.
This constraint is even weaker than the requirement that mutations should be naturalistically plausible. On one hand the idea of mutations being limited (in some specified way) is vague and not very useful. However it does show how easy it is to avoid the accusation that Universal Darwinism is empty and vacuous. Just put a limit (any limit) on the type of mutations permitted by the theory, and you have a falsifiable theory.
If the Darwinist theory is the proper metatheoretical model of all social sciences including economic science, as the authors of this book claim, what does it tell us economists about the maximization hypothesis? In regard with the consumption of matter and energy of the living entities the authors of Darwin’s Conjecture asserts that they “have limited capacities to absorb” (p.33). However, there is a magnitude which is maximized by replicators - the number of offspring. But enough disappointing, human reality seems to be depicted by an upside down situation: it is rather the number of offspring which humans strive to limit and it is each individual’s consumption of matter and energy that is maximized. The authors do not seem aware of this serious contradiction and are not at all concerned to account for by a more comprehensive principle that reconcile these two principles of limitation and maximization, which apply differently to human and non-human living worlds.
I think this doesn't really hit the mark. Human offspring is what a naive human-centric, DNA-centric view of evolution might point to as evolution's maximand. It isn't that far from the mark - as today's 7 billion humans attests to. However, human DNA is only one of the things copied in evolution. There are also plants, bacteria and memes. Memes are what explains the demographic transition which Paul alludes to. Gene-meme coevolution is the only credible theory that accounts for the reduced levels of offspring creation associated with being a rich westerner. There's been quite a bit of effort put into understanding how it works.
There is no "principle of limitation" that only applies to the human world. Maximization explains things pretty neatly. The maximand isn't human offspring, though - memes count too. We are already witnessing the effects of high meme concentrations on human fertility - for example, in Japan. In the future, the number of humans may well dwindle - as existing humans get sucked into cyberspace and fail to procreate. This would not be a failure of Darwinian evolution - it will just illustrate one type of information being copied at the expense of another. Ultimately, entropy is the maximand - as has been argued by many.
Entropy maximization is consistent with Paul's claim that we should look to physics - and not biology - for the answer here. However, entropy maximization isn't really a replacement for evolutionary theory - it is more of a useful adjunct.
 We have previously seen that copying is found ubiquitously in nature, from spreading ripples to propagating cracks, from growing crystals to scattering radiation. Of course, copying, variation and selection are the basis of Darwinian evolutionary theory. The copying can take a variety of forms, but the most basic is positional inheritance.
We have previously seen that copying is found ubiquitously in nature, from spreading ripples to propagating cracks, from growing crystals to scattering radiation. Of course, copying, variation and selection are the basis of Darwinian evolutionary theory. The copying can take a variety of forms, but the most basic is positional inheritance. The products of positional inheritance often form tree-like structures. The roots and branches of plants resemble trees - and actually are phylogenetic trees of plant cells, laid down in order during development - in a combination of phylogeny and ontogeny. Similarly, lightning, propagating cracks, fractal drainage patterns, and crystalline dendrites are all associated with prominent visual trees. In each case, these are family trees, that show the path of descent. That these trees are in fact family trees can often be easily verified by filming their formation in slow motion. Videos of lightning strikes slowed down show that forks always descend from existing branches. If you look at videos of bullets hitting panes of glass you will see that propagating cracks behave in a similar manner - the cracks spread outwards in a radial pattern where each crack descends from an earlier parent crack.
The products of positional inheritance often form tree-like structures. The roots and branches of plants resemble trees - and actually are phylogenetic trees of plant cells, laid down in order during development - in a combination of phylogeny and ontogeny. Similarly, lightning, propagating cracks, fractal drainage patterns, and crystalline dendrites are all associated with prominent visual trees. In each case, these are family trees, that show the path of descent. That these trees are in fact family trees can often be easily verified by filming their formation in slow motion. Videos of lightning strikes slowed down show that forks always descend from existing branches. If you look at videos of bullets hitting panes of glass you will see that propagating cracks behave in a similar manner - the cracks spread outwards in a radial pattern where each crack descends from an earlier parent crack. One thing that evolving systems typically need, in order to exhibit complex adaptations, is high-fidelity copying. Excessive noise often results in inherited information getting lost - and this leads to the disintegration of complex adaptations. However, positional inheritance often has pretty high fidelity - allowing adaptations based on it to remain stable. If you think of it in terms of coordinates in the universe that are unknown to an observer, by learning the location of an object an observer gains a considerable quantity of information - that object's coordinates in three dimensional space. If the offspring is within 1 meter of the parent, then that's about 265 bits of mutual information copied with high-fidelity. Say 350 bits of spacetime. Of course in practice, few positional inheritance systems take up the whole universe, so 350 bits is an upper limit. 350 bits is peanuts compared to biological systems, but it represents a search space of considerable size - it's enough for some non-trivial optimization processes to take place.
One thing that evolving systems typically need, in order to exhibit complex adaptations, is high-fidelity copying. Excessive noise often results in inherited information getting lost - and this leads to the disintegration of complex adaptations. However, positional inheritance often has pretty high fidelity - allowing adaptations based on it to remain stable. If you think of it in terms of coordinates in the universe that are unknown to an observer, by learning the location of an object an observer gains a considerable quantity of information - that object's coordinates in three dimensional space. If the offspring is within 1 meter of the parent, then that's about 265 bits of mutual information copied with high-fidelity. Say 350 bits of spacetime. Of course in practice, few positional inheritance systems take up the whole universe, so 350 bits is an upper limit. 350 bits is peanuts compared to biological systems, but it represents a search space of considerable size - it's enough for some non-trivial optimization processes to take place. Positional inheritance doesn't lead to adaptations on the scale seen in biological evolution. Various problems and limitations reduce its scope for generating adaptations. Many inorganic systems exhibiting positional inheritance lack important properties found in the organic domain.
Positional inheritance doesn't lead to adaptations on the scale seen in biological evolution. Various problems and limitations reduce its scope for generating adaptations. Many inorganic systems exhibiting positional inheritance lack important properties found in the organic domain.

 The term "refactoring" commonly refers to a set of useful computer programming techniques which involve making structural changes to a program without altering its function. Refactoring is often done before making functional changes. If you launch into making changes without some preliminary refactoring, you are often more likely to make mistakes. Refactoring frequently allows you to make changes without breaking tests - so you can verify that your changes don't break anything. If you need to make a bunch of changes, it sometimes helps to make ones that you can verify do no damage first.
The term "refactoring" commonly refers to a set of useful computer programming techniques which involve making structural changes to a program without altering its function. Refactoring is often done before making functional changes. If you launch into making changes without some preliminary refactoring, you are often more likely to make mistakes. Refactoring frequently allows you to make changes without breaking tests - so you can verify that your changes don't break anything. If you need to make a bunch of changes, it sometimes helps to make ones that you can verify do no damage first.
 I came up with the idea of branch tip evolution in 2012 by thinking about
I came up with the idea of branch tip evolution in 2012 by thinking about 



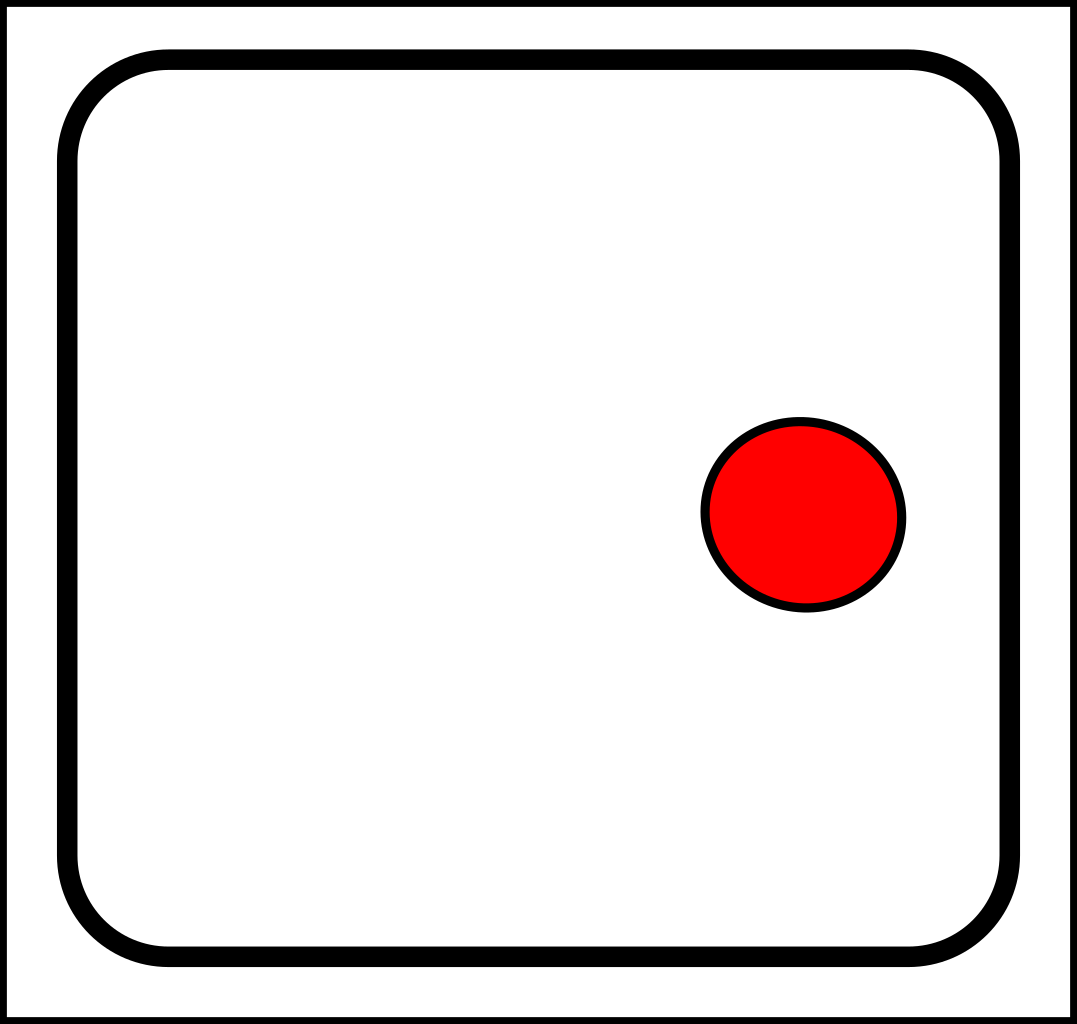
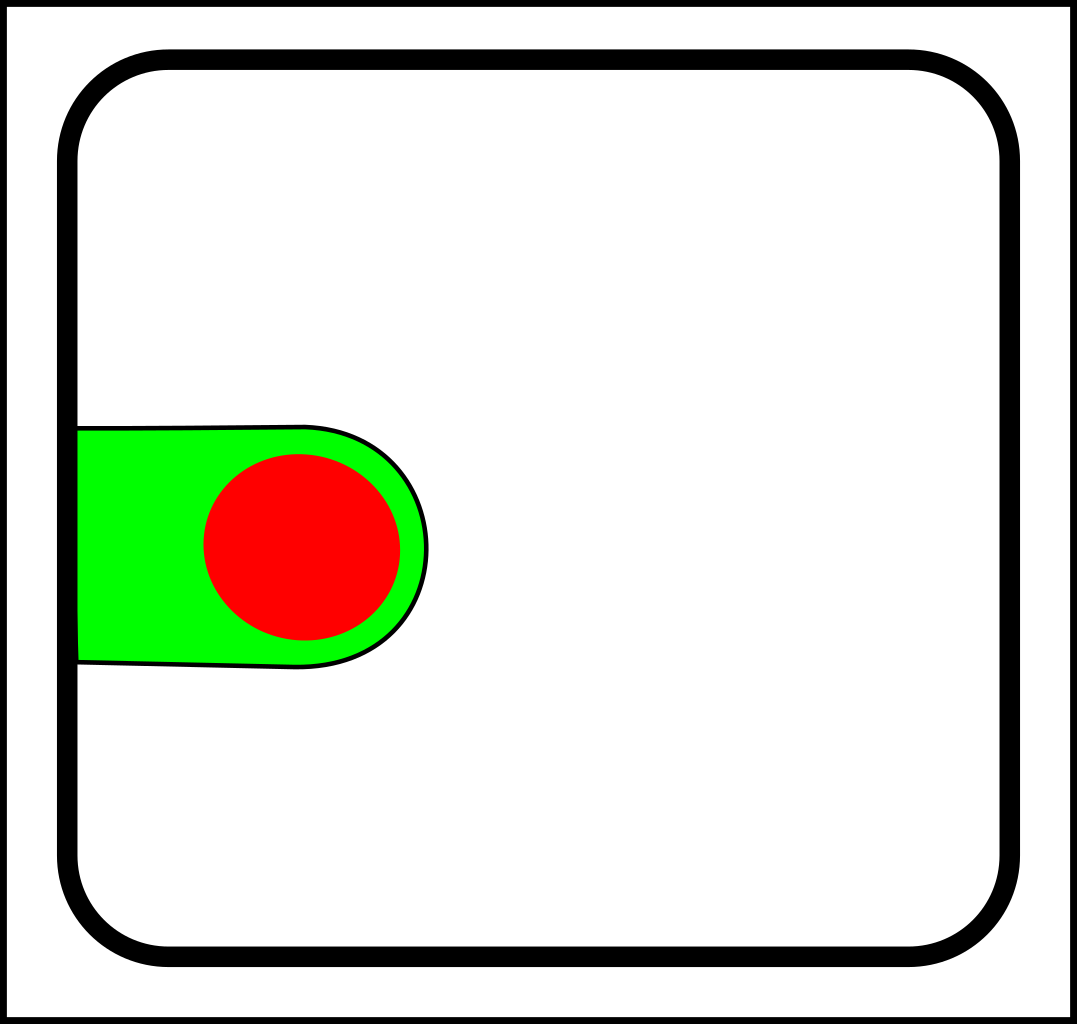

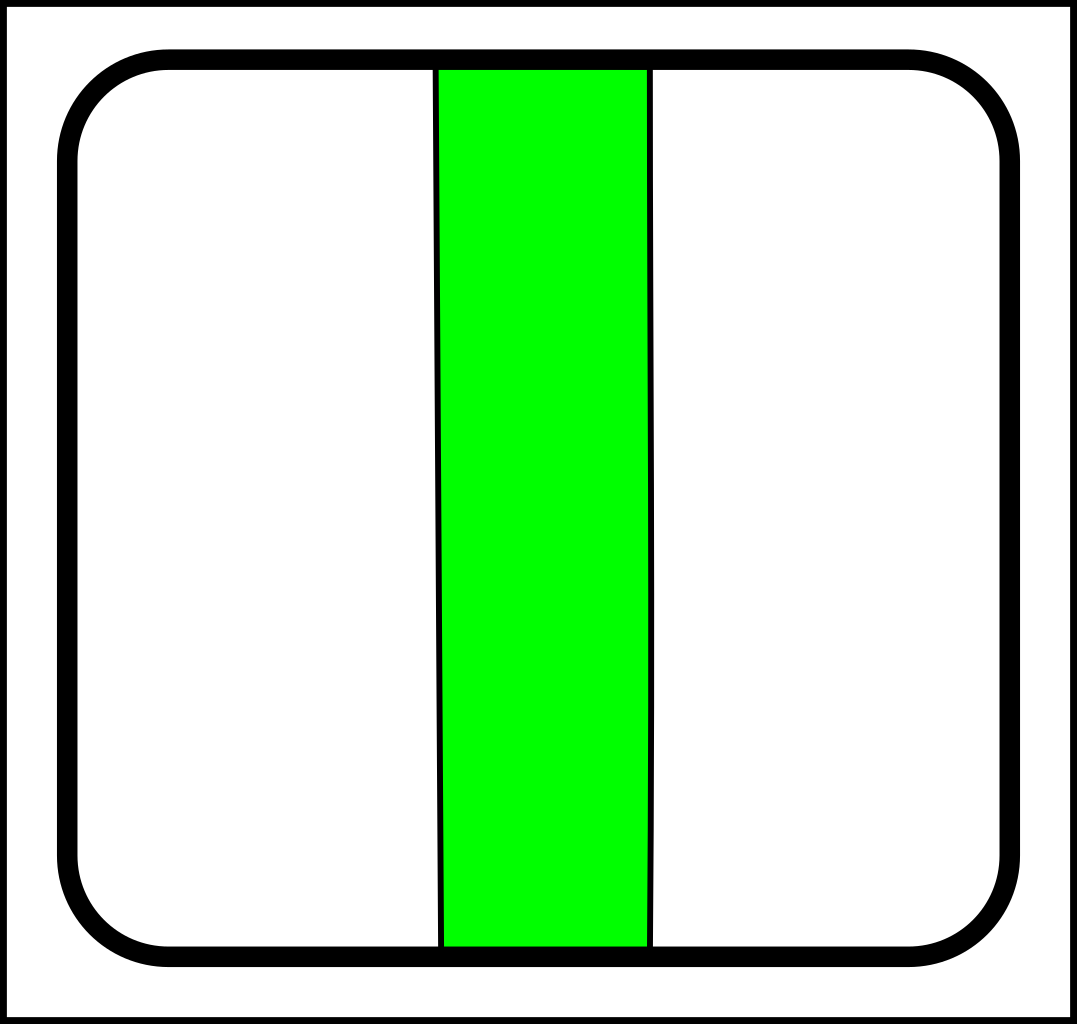
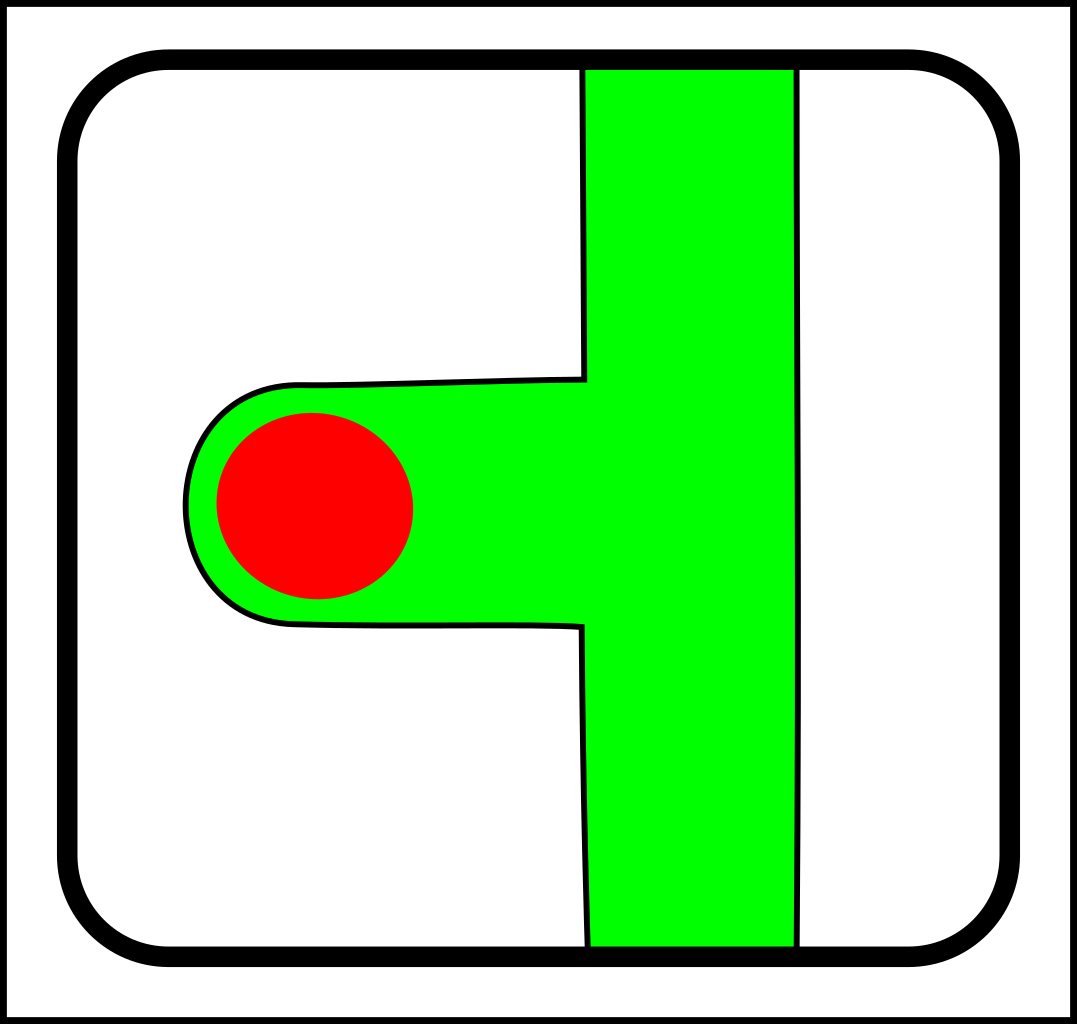


 I just noticed that Addy Pross offered a criticism of universal Darwinism
I just noticed that Addy Pross offered a criticism of universal Darwinism  Tree-shaped patterns in nature are frequently found in both the organic and inorganic domains and
demand a common explanation. One type of explanation is a functional one. Many trees
efficiently drain basins. Electrical discharges can be modeled as draining electrical
charge and propagating cracks do something similar with stress. The maximum entropy production
principle generalizes this idea so that it can be applied to evolutionary family trees
as well as to physical trees.
Tree-shaped patterns in nature are frequently found in both the organic and inorganic domains and
demand a common explanation. One type of explanation is a functional one. Many trees
efficiently drain basins. Electrical discharges can be modeled as draining electrical
charge and propagating cracks do something similar with stress. The maximum entropy production
principle generalizes this idea so that it can be applied to evolutionary family trees
as well as to physical trees.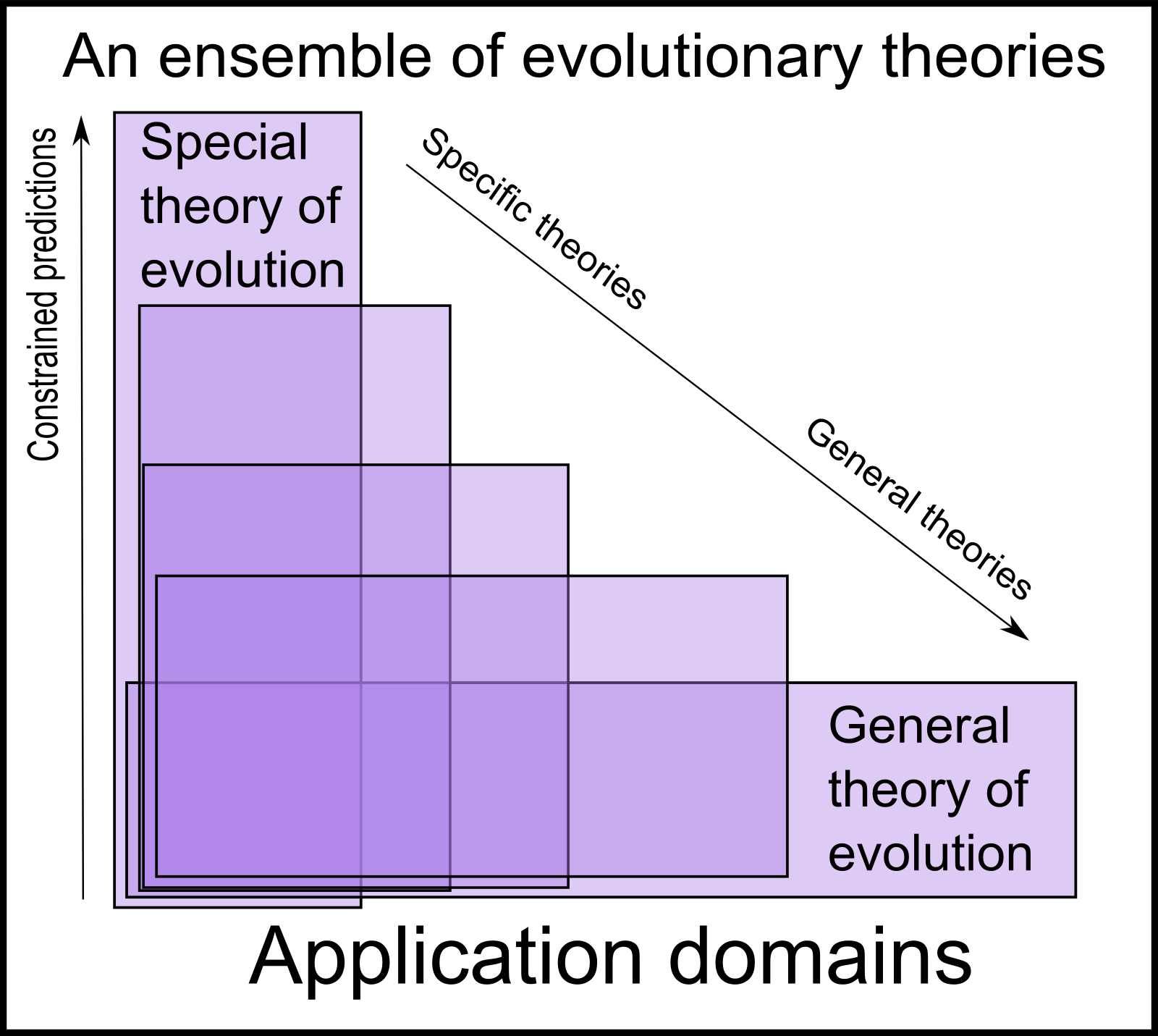
 It's well known that Donald Campbell was an early pioneer of cultural evolution. Most in the field acknowledge his work as influential or historically important. One of his early papers on "Blind variation and selective retention" dates from 1960. Later in life, Donald Campbell explored the limits of evolutionary theory - applying it to a range of inorganic phenomena. In particular, there's the following paper:
It's well known that Donald Campbell was an early pioneer of cultural evolution. Most in the field acknowledge his work as influential or historically important. One of his early papers on "Blind variation and selective retention" dates from 1960. Later in life, Donald Campbell explored the limits of evolutionary theory - applying it to a range of inorganic phenomena. In particular, there's the following paper: Computer programmers have a proverb that goes:
Computer programmers have a proverb that goes:
 One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, fit offspring should be ancestral to fit descendants. Without this condition being met, adaptations can't get off the ground.
One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, fit offspring should be ancestral to fit descendants. Without this condition being met, adaptations can't get off the ground. In 2011, Paul Fudulu has written a somewhat-interesting critique of Universal Darwinism - titled:
In 2011, Paul Fudulu has written a somewhat-interesting critique of Universal Darwinism - titled: 













