 The concept of "selection" is at the heart of evolutionary theory. Some people assign that role to natural selection - but the main difference between selection and natural selection seems to be the idea that selection also includes artificial selection. However, the whole idea that selection by man is somehow "unnatural" doesn't deserve much in the way of scientific respect.
The concept of "selection" is at the heart of evolutionary theory. Some people assign that role to natural selection - but the main difference between selection and natural selection seems to be the idea that selection also includes artificial selection. However, the whole idea that selection by man is somehow "unnatural" doesn't deserve much in the way of scientific respect.The definition of "selection" in a scientific context is the subject of some controversy. To give an example, Hull, Langman and Glenn (1999) define 'selection' as follows:
we define selection as repeated cycles of replication, variation and environmental interaction so structured that environmental interaction causes replication to be differential.
For me, such definitions are problematical. "Selection" is an ordinary English word, which means "a choice from alternatives". Choosing from alternatives strikes me as being a simple and sensible scientific concept. However, the idea of structured "repeated cycles of replication, variation and environmental interaction" is long-winded and complex. I generally favour simple scientific concepts on basic grounds of parsimony.
It is quite common for definitions of natural selection to be very specific. For example, they often mention "populations" or "inherited traits". Being overly specific is a common problem in science. Scientists favour general theories that explain lots of data over more specific ones that don't explain so much. However while inductive inference proceeds from specific examples to general theories, it sometimes takes a while for it to get there. Selection takes place between alternatives. The term "population" carries with it a lot of irrelevant and unnecessary baggage. As for selection involving "inherited traits" - that excludes selection that acts on traits that are not inherited. Yet much the same theory applies to these traits. Defining selection - or natural selection - as involving "inheritance" is a narrow and blinkered approach to the topic.
The idea of choosing from alternatives is really a very general one. If you think about it, practically any form of change can be described as being the result of a selection between different types of possible change. Selection is often contrasted with mutation - but even mutations can be described as a form of choice - a choice between different possible types of mutation. In evolutionary biology selection of which organisms die, which organisms reproduce and which organisms are chosen as mates are considered to be important forms of selection - while selection of which mutations take place and which do not is not generally considered to be a form of "selection" at all. The "selection" criteria for mutations is often assumed to be pretty trivial: mutations are chosen "at random".
Does the generality of the concept of selection make the concept any less scientific? If practically any change can be explained in terms of selection, then isn't invoking selection as an explanation vacuous? Not really: scientific explanations usually say what type of entities are being selected. An explanation invoking selection on organisms can be perfectly scientific and testable. Specifying the entities being selected in a theory rules out a wide range of other selective explanations. Also, an explanation in terms of selection is always possible - but it might not be likely: other explanations might be more parsimonious. Compare with information. You can describe anything in terms of information. However we still have useful theories associated with information. Being very general is not necessarily a fatal flaw.
In biology, selection is the primary explanation of adaptation. However, goodness of fit is invoked in other areas of science - and very often selection is involved in its creation. For example, observation selection explains goodness of fit between observers and the universe they observe.
There's a grand thesis associated with selection in which selection explains most goodness of fit. Gary Cziko and Donald Campbell are among the proponents of this view. For example, they describe learning strategies as either "instructional" or "selectionist" and then argue that what looks like instructional learning often turns out to be selectionist. This view also has its critics - who describe the act of labeling "instructional" learning as being really "selectionist" as obtuse. For example, Henry Plotkin raises this objection in Evolutionary Worlds Without End.
My sympathies lie with Cziko and Campbell on this issue - though I have some issues with their presentation. It's better to have a broad concept of selection - and this leads naturally to a grand unified theory of selection in which selection explains most goodness of fit. The idea means that selection must be involved in all template copying. For example, the "goodness of fit" between a footprint in the sand and the corresponding foot must be explicable in terms of selection acting against the grains of sand that were displaced. This is a somewhat counter-intuitive use of the term "selection" - but it perfectly possible to imagine the foot as the selecting agent and the grains of sand as being the selectees.
The existence of goodness of fit usually means that - at some stage a selector has acted on alternative choices - and those remaining 'fit' the selector. Of course goodness of fit by chance is always a possibility. However it is a possibility that is usually easily quantified and rejected. Almost all goodness of fit in nature can be attributed to selection.
 Fitness landscapes are a common way of visualizing the relationship between variables associated with an organism
and fitness. The height of the landscape represents fitness and the domain over which the fitness landscape is defined
is composed of other variables associated with the organism. Most frequently the information associated with heritable
elements is used - but other variables affecting fitness could be included as well - such as environmental factors.
Fitness landscapes are a common way of visualizing the relationship between variables associated with an organism
and fitness. The height of the landscape represents fitness and the domain over which the fitness landscape is defined
is composed of other variables associated with the organism. Most frequently the information associated with heritable
elements is used - but other variables affecting fitness could be included as well - such as environmental factors.
 In evolutionary processes, sometimes parents and offspring are indistinguishable and other times, they are very different.
In evolutionary processes, sometimes parents and offspring are indistinguishable and other times, they are very different. One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, fit offspring should be ancestral to fit descendants. Without this condition being met, adaptations can't get off the ground.
One of the commonly-specified requirements for Darwinian systems is that fitness must be heritable. In other words, on average, fit offspring should be ancestral to fit descendants. Without this condition being met, adaptations can't get off the ground.
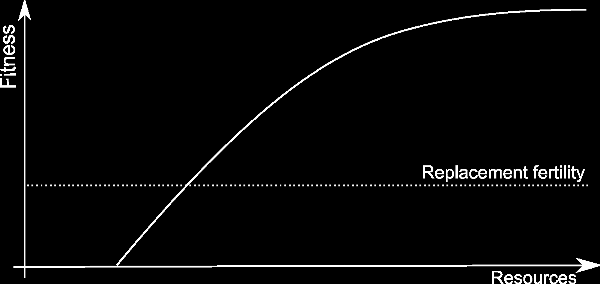 Organisms can covert resources into fitness. However they face limits when doing so: too few resources and they can't even live, let alone reproduce; too many resources and the organism can't utilize them fast enough. This situation is illustrated on graph to the right:
Organisms can covert resources into fitness. However they face limits when doing so: too few resources and they can't even live, let alone reproduce; too many resources and the organism can't utilize them fast enough. This situation is illustrated on graph to the right:













